Генетика популяцій
Статевий процес (зачатки якого реалізуються навіть у прокаріотів, див. розділ 5) створює умови обміну генами між особинами одного виду, що проживають на спільній території. Саме така група особин, яка узагальнює певну сукупність генів, - популяція - є елементарною одиницею еволюційного процесу.
Популяцією називають групу особин одного виду, що мають спільний ареал (мешкають на спільній території) і спільний генофонд, відокремлений від сусідніх груп (рис. 8.1). Між особинами популяції реалізуються тісні генетичні відносини - більш-менш вільне схрещування в межах групи. Відокремлення від сусідніх популяцій здійснюється внаслідок певної обмеженості таких відносин із іншими групами (ізоляції від інших популяцій). Еволюційні зміни виду відбуваються шляхом зміни генофонду - загальної сукупності генів - кожної такої популяції.

Популяція не є просто арифметичною сукупністю особин, це певна система, якій властиві свої особливості структури та функціонування. Особливості популяції як системи базуються на видових та індивідуальних властивостях особин, що входять до її складу, але не можуть бути зведені до суми цих властивостей. З іншого боку, властивості організмів, які становлять популяцію, залежать від особливостей цієї популяції та формуються в умовах її існування. Отже, популяційна генетика, предметом якої є генетичні параметри груп особин і зміна цих параметрів, відрізняється певною специфікою проблематики та методів досліджень.
До найважливіших параметрів, що характеризують популяцію, відносять чисельність, мінливість, структурованість і особливості розмноження (системи схрещування), а також частоти генів (алелів) та генотипів. Ці параметри разом з іншими зумовлюють унікальність генофонду популяції та її генетичну структуру.
У найпростішому випадку, без урахування міграційних процесів, чисельність популяції залежить від співвідношення двох величин -народжуваності та смертності. Якщо народжуваність (кількість новонароджених, віднесена на одну особину, за певних час t) позначити як a, смертність (кількість смертей на одну особину за той самий час) -як b, то різниця між ними r дасть уяву про зміну чисельності популяції протягом дослідженого періоду:
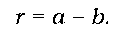
На честь Мальтуса (Thomas Robert Malthus), який першим звернув увагу на різну швидкість росту населення та необхідних ресурсів, величину r називають мальтузіанським параметром. Використовуючи цей параметр, миттєву швидкість зміни чисельності популяції N (зміну чисельності за певний момент часу, коли тривалість досліджуваного періоду t наближається до нуля) можна виразити рівнянням
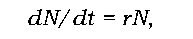
розв'язання якого дає експоненціальний характер зростання чисельності в часі (швидкість є тим вищою, чим більшою є чисельність популяції).
Ця експоненціальна м,одель росту популяції враховує лише два параметри - народжуваність і смертність. Вона описує вибухоподібне
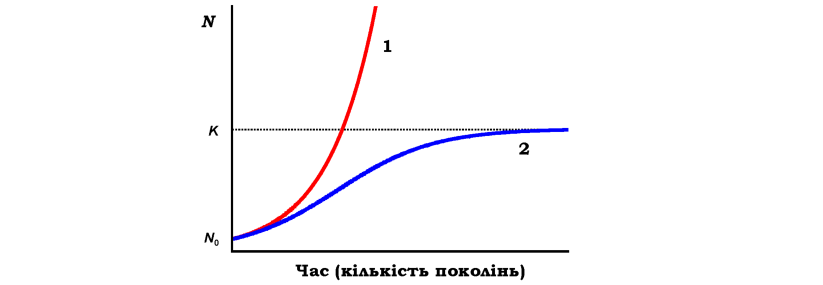
зростання чисельності в умовах відсутності тиску середовища й наявності нескінченно великих ресурсів (рис. 8.2).
 |
|
Рис. 8.2. Дві криві росту чисельності популяції N: експоненціальна (1) і логістична (2). N0 - вихідна чисельність популяції, K - верхня критична межа чисельності популяції, яка залежить від ресурсів середовища |
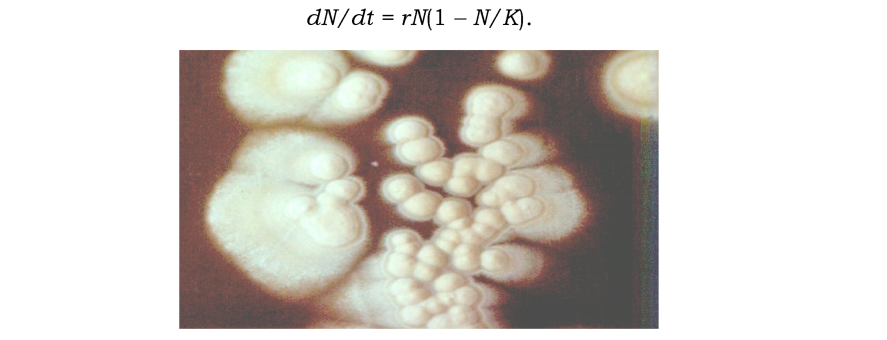
Обмеженість ресурсів зумовлює певний "тиск середовища" на здатність популяції до експоненціального росту (див. рис. 8.3). Тобто існує верхня критична межа чисельності популяції K, яка визначається спроможністю середовища підтримувати на обмеженому ареалі існування лише обмежену кількість особин одного виду. Тоді швидкість зростання популяції залежатиме від того, наскільки в даний момент чисельність N наблизилась до K, (логістична модель росту популяції, рис. 8.2):
 |
|
Рис. 8.3. Зростання колоній дріжджів на поверхні живильного середовища в чашці Петрі. Добре помітно сповільнення росту колоній у напрямку найближчих сусідніх колоній і більш інтенсивний ріст у протилежному. Різницю темпів росту можна пояснити зменшеною концентрацією поживних речовин (ресурси середовища) і підвищеною концентрацією метаболітів у зоні сусідства |
Чисельність популяції та її динаміка є вагомими показниками стану популяції. Не менш важливим параметром є так звана ефективна чисельність популяції (Ne). Справа в тому, що не всі особини, які здатні залишати нащадків, насправді роблять репродуктивний внесок у відтворення популяції. У найпростішому випадку Ne можна визначити як чисельність особин, які залишають потомство. Якщо чисельності самців Nm і самок Nf приблизно однакові (вважаючи, що всі самці й самки є репродуктивно спроможними), то ефективна чисельність популяції просто дорівнює їхній сумі:
Залежність ефективної чисельності популяції від співвідношення статей для різних груп з однаковим кількісним складом показано в табл. 8.1. Очевидно, що ця величина дорівнює сумі самців і самок репродуктивного віку лише за умови їхньої однакової чисельності (останнє рівняння зводиться до попереднього, якщо Nm = Nf). У всіх інших випадках Ne< (Nm + Nf). На величину Ne впливає також розмір нерепродуктивної частини популяції (різні вікові групи або "робочі особини" у бджіл чи мурашок), неоднакова репродуктивна здатність, коливання загального кількісного складу популяції та ряд інших факторів.
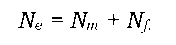
У випадку значної різниці між чисельністю самців і самок ефективну чисельність популяції знаходять за виразом
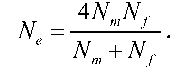
|
Таблиця 8.1. Теоретично розрахована ефективна чисельність популяції Neдля групи зі 100 особин при різних співвідношеннях самців Nmі самок Nf |
||||||||||||||||||||||||||||||||
|
Наявність відмін між особинами одного виду є необхідною умовою еволюційних змін популяції. Мінливість фенотипів може бути викликана факторами середовища (неспадкова мінливість), генетичними відмінами (спадкова мінливість) або ж обома факторами, як це характерно для кількісних ознак.
Мінливість кількісних ознак при популяційних дослідженнях характеризують середнім значенням ознаки, дисперсією, а також коефіцієнтами варіації та успадкування (див. розділ 3). Мінливість альтернативних (якісних) ознак визначають часткою в популяції певної форми (або морфи) цієї ознаки - частотою фенотипу. Основними показниками генетичної мінливості в популяції є частоти генів і генотипів. Рівень генетичної мінливості популяції є основним джерелом для її потенційної адаптивної зміни.
Якщо в популяції існує всього один алель гена, його називають мономорфним, якщо два й більше - поліморфним, а наявність у популяції кількох алельних форм гена - генетичним поліморфізмом. Частка поліморфних генів серед усіх проаналізованих - поліморфність (Р -є одним із показників генетичної мінливості популяцій. Умовним критерієм поліморфності гена є частота його алелів: ген вважають поліморфним, якщо частки хоча б двох алелів перевищують 0,05 або 0,01 (два пороги поліморфності). Якщо ж ця частота не перевищує порого-ве значення, алель відносять до рідких алелів або до мутацій. Приклад обчислення поліморфності Р наведено в табл. 8.2.
|
Таблиця 8.2. Обчислення середньої поліморфності (P) чотирьох популяцій |
||||||||||||||||||||||||||
|
||||||||||||||||||||||||||
Ще одним показником генетичної мінливості популяції є середня гетерозиготність (H). У кожної особини в популяції певна частина генів перебуває в гетерозиготному стані. Частка таких генів (від проаналізованих) характеризує гетерозиготність цієї особини. Усереднена величина індивідуальних значень гетерозиготності для всіх обстежених особин є середньою гетерозиготністю популяції H .
Гетерозиготність значною мірою залежить від частоти алелів. Якщо частоти двох алелів одного гена дорівнюють 0,9 та 0,1, то частка гетерозигот становить 2 х 0,9 х 0,1 = 0,18 (див. розділ 3 і рівняння Харді -Вайнберга нижче). За частот двох алелів 0,4 і 0,6 частка гетерозигот становитиме вже 2 х 0,4 х 0,6 = 0,48. Найвищими значення гетерозиготності будуть при однаковості частот алелів кожного з генів (1/2 і 1/2 для двоалельних генів; 1/3, 1/3 і 1/3 для триалельних і т. п.).
Структурованість означає наявність у популяції окремих груп; у межах групи особини об'єднані між собою за якимось критерієм. Розрізняють вікову, статеву, просторову, екологічну та генетичну структуру популяції.
Співвідношення в популяції осіб різного віку й статі визначає її вікову та статеву структуру відповідно. Просторова структура - це характер розподілу на популяційному ареалі окремих особин та їхніх угруповань. Як приклади можна назвати острівний і гомогенний розподіли. Екологічною структурою називають залежне від біотичних і абіотичних факторів середовища розподілення популяції на групи.
Подібна структурованість визначає особливості генетичної структури популяції (див. нижче) і формування генофонду наступних поколінь.
Із двох загальних типів розмноження - статевого та нестатевого -у популяційній генетиці основну увагу приділяють саме статевому способу розмноження, який забезпечує суттєвий рівень генетичної мінливості за рахунок рекомбінацій. Усе різноманіття видів статевого розмноження можна розділити на кілька типів.
Панміксія означає однакову ймовірність мати спільне потомство для будь-якої пари осіб протилежної статі. При цьому відсутня будь-яка вибірковість. Наявність такої вибірковості має місце при різного роду шлюбній асортативності - переважному формуванню шлюбних пар певного типу. Крайніми випадками асортативності є аутб-ридинг та інбридинг.
Аутбридинг - схрещування найбільш генетично віддалених партнерів - ліній, підвидів, або в окремих випадках - видів чи навіть родів. Останні два типи схрещувань ще називають міжвидовою чи міжродовою гібридизацією. Схрещування між генетично близькими формами називають інбридингом. Це, як правило, схрещування в межах однієї генетичної лінії, між родичами, самозапліднення (наприклад, поширене серед рослин самозапилення).
ГЕНЕТИЧНА СТРУКТУРА ПОПУЛЯЦІЇ. ЗАКОН ХАРДІ -ВАЙНБЕРГА
Під генетичною структурою популяції розуміють співвідношення особин із різними генотипами, особливості формування генетичних зв'язків (систему схрещувань) і розподіленість популяцій на ряд угрупувань (субпопуляцій), пов'язаних між собою потоками алелів. Основними параметрами генетичної структури популяцій є частоти генів і генотипів.
У популяції може існувати один, два, три чи більше (не обмежено) алелів певного гена. У конкретної диплоїдної особини з популяції -два алелі (однакових у гомозигот і різних у гетерозигот) для аутосом-них генів. Гени статевих хромосом при XY-типі визначення статі представлені одним алелем у самців і двома - у самок; при ZW-типі -навпаки (див. розділ 6).
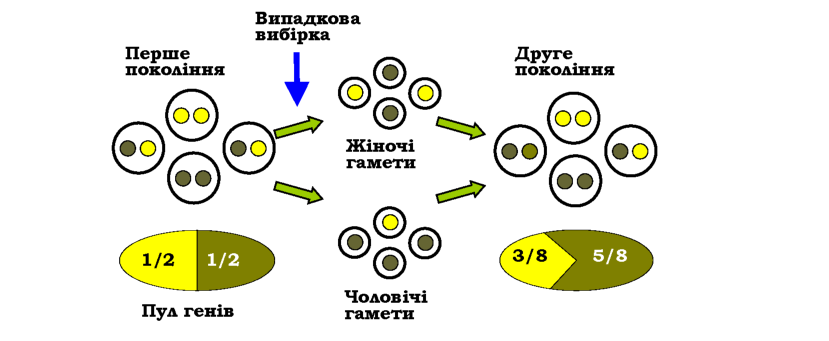
Частота алеля визначається як відношення кількості копій даного алеля до загальної кількості алелів цього гена в усіх особин популяції. Якщо в популяції існує два алелі певного гена (скажімо, А та а), їхню частоту можна позначити як рл і qa, або просто p і q. Частота генотипу - це частка особин із певним генотипом у популяції, яку можна позначити як /(ЛА), /(Ла), /(аа).
У 1908 р. Xарді та Вайнберг (Godfrey Hardy, Wilhelm Weinberg) незалежно один від одного дійшли висновку, що за певних умов менделівсь-кий механізм спадкування забезпечує постійність (із покоління в покоління) співвідношення генотипів у популяції для будь-яких частот алелів.
Розглянемо приклад. Група із N = 200 особин має такий склад:
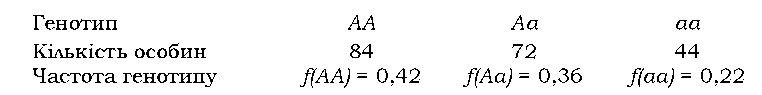
|
Знайдемо частоту домінантного алеля. Гомозиготи мають по два однакових алеля, гетерозиготи - лише один; загальна кількість алелів у популяції диплоїдів дорівнює подвоєній чисельності особин. Маємо |
 |
Розрахунок частот алелів дасть знову 0,6 та 0,4, і в подальших поколіннях ані ці частоти алелів, ані частоти генотипів змінюватись не будуть. Таке співвідношення генотипів у популяції, яке здатне автоматично зберігатись протягом нескінченно великої кількості поколінь, називають рівноважним співвідношенням генотипів, а явище підтримання постійного співвідношення генотипів протягом поколінь -
генетичною рівновагою. У розглянутому прикладі частоти генотипів вихідного покоління не перебували в стані рівноваги, але перейшли до рівноважного стану після першого ж панміктичного схрещування.
Отже, співвідношення генотипів у популяції буде постійним протягом нескінченної кількості поколінь для будь-яких частот алелів. Ця закономірність, відома як закон Харді - Вайнберга, описує ключову особливість популяцій - здатність до підтримання сталості частот генотипів (генетичної рівноваги). Закон Харді - Вайнберга виконується тільки для ідеальних (менделівських) популяцій: нескінченно великих панміктичних популяції диплоїдного виду зі статевим розмноженням, при однаковій життєздатності всіх генотипів і відсутності інших факторів динаміки популяції - факторів, які змінюють частоти генотипів і/або алелів (див. нижче).
Рівноважні співвідношення частот генотипів задаються піднесенням до квадрата суми частот алелів. Для двох алелів рівняння Харді -Вайнберга має такий вигляд:
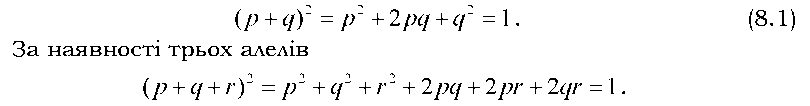
Якщо в популяції (чи в групі особин) співвідношення генотипів не рівноважне (тобто реальні співвідношення частот генотипів не відповідають теоретично очікуваним на основі закону Харді - Вайнберга для даних частот алелів), популяція перейде до стану генетичної рівноваги після першого ж панміктичного схрещування (як у розглянутому вище прикладі). Це справедливо для генів, які містяться в аутосомах.
Розглянемо встановлення генетичної рівноваги для генів, зчеплених зі статтю, зокрема з Х-хромосомою при ХY-механізмі визначення статі. У цьому випадку самки (гомогаметна стать) матимуть 2/3 всіх наявних у популяції генів (розташованих у Х-хромосомі), а самці - 1/3. У дочірньому поколінні завдяки крис-кросному механізму передачі статевих хромосом самці матимуть усі свої Х-хромосоми від матерів, а самки - одну від матері, а другу від батька. Тоді, якщо частоти алелів не однакові в самців і самок, то в кожному наступному поколінні в самців частота алеля буде дорівнювати частоті алеля самок попереднього покоління, а в самок - середньому арифметичному частот алелів самців і самок попереднього покоління. Оскільки самки мають 2/3 від усієї кількості алелів (Х-хромосом) у популяції, а самці - 1/3, то рівноважні значення частоти алелів такі:
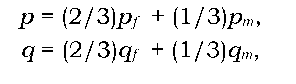
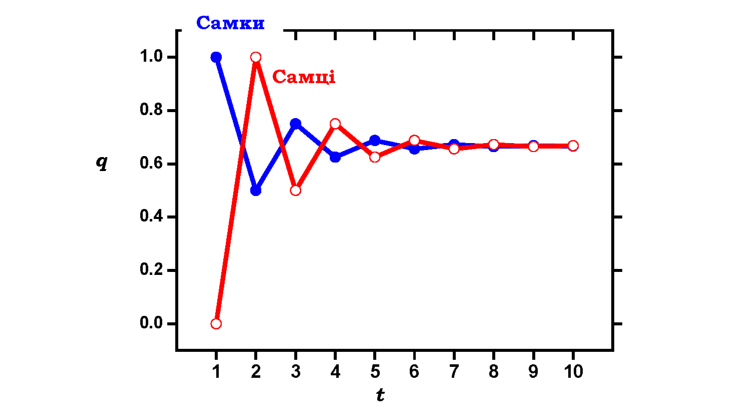
де індекс f указує на частоту (рівноважну чи нерівноважну) відповідного алеля в самок, m - у самців. У стані генетичної рівноваги частоти алелів у самок і самців однакові. Для прикладу, розглянутого на рис. 8.4, рівноважним значенням частоти алеля буде q = (2/3)qf + (1/3)qm = 2/3. Важливо розуміти, що при встановленні генетичної рівноваги за Х-зчепленими генами, частоти алелів змінюються в межах кожної статі, залишаючись постійними для популяції в цілому.
 |
|
Рис. 8.4. Установлення генетичної рівноваги для Х-зчеплених генів. По осі абсцис - номер покоління t, по осі ординат - частота алеля q. Вихідні значення частоти алеля для самців qm = 0, для самок qf = 1. З кожним поколінням різниця між qm і qf зменшується, а частота алеля наближається до рівноважного значення 2/3 |
Перевірка рівноважності популяції проводиться шляхом порівняння (за допомогою методу %2, див. розділ 3) реального розподілу генотипів з очікуваними частотами генотипів, визначеними відповідно до закону Харді - Вайнберга. Така перевірка є можливою лише тоді, коли можна експериментально оцінити частоту гетерозигот, а саме при кодомінуванні.
За повного домінування гомозиготи АА та гетерозиготи Аа відносять до одного й того ж фенотипу, тому шляхом безпосереднього підрахунку неможливо оцінити ані частоту гетерозигот, ані частоту алелів. Це можна зробити на основі закону Харді - Вайнберга, припустивши наявність у популяції генетичної рівноваги. Зрозуміло, що в даному випадку реальні значення частот алелів можуть бути далекими від одержаних гіпотетичних.
ФАКТОРИ ДИНАМІКИ ГЕНЕТИЧНОЇ структури ПОПУЛЯЦІЙ
Нагадаємо, що умовами виконання закону Харді - Вайнберга є:
• необмеженість чисельності популяції;
• панміксія;
• відсутність мутацій;
• відсутність міграцій;
• відсутність добору.
У природі практично не буває популяцій, для яких виконувалися б усі ці умови. Якщо існують суттєві відхилення від них, то залежно від ситуації можуть змінюватись частоти генотипів при збереженні частот алелів, або ж будуть змінюватись як частоти генотипів, так і частоти алелів.
Слід, проте, зауважити, що в стаціонарних умовах середовища та для достатньо великих популяцій рівняння 8.1 виявляється справедливим для більшості генів: велика кількість генів не є об'єктами добору; для багатьох генів розподіл алелів і генотипів є подібним для різних популяцій, що змішуються між собою під час міграцій.
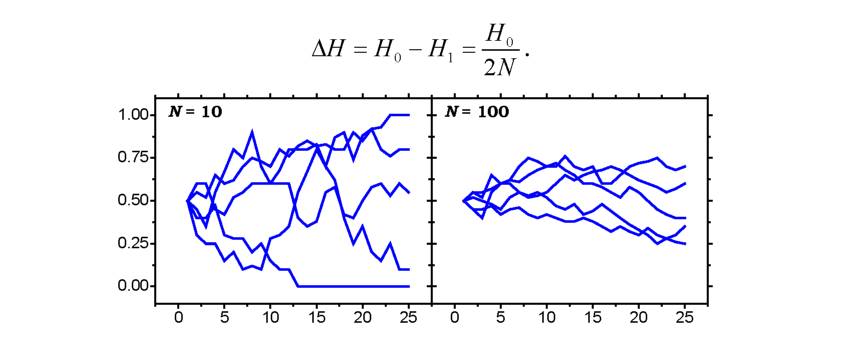
Чисельність популяцій завжди є величиною скінченною. Якщо (у разі відсутності добору, мутацій і міграцій) популяція характеризується незначною величиною Ne, то при формуванні пулу гамет і генофонду наступної генерації значно підвищується ймовірність випадкових відхилень від середньої частоти того чи іншого алеля. Процес ненаправленої зміни частот алелів у малих популяціях під впливом випадкових факторів називають дрейфом генів.
Ці випадкові коливання спричинюють непередбачені зміни частот генів у ряду генерацій (рис. 8.5, 8.6). Кінцевим результатом цього процесу буде елімінація алеля з популяції або його фіксація (частота алеля фіксується на 100 % рівні), причому тим швидше, чим меншим є значення Ne.
 |
|
Рис. 8.5. Процес випадкової зміни генних частот протягом однієї генерації в популяції, що складається з чотирьох гермафродитних особин, при репродукції яких відбувається випадкове об'єднання гамет (за Кімура, 1985) |
Випадковість і непередбаченість змін генетичної структури конкретної популяції зумовлює використання ймовірністного підходу, лише за допомогою якого і є можливим вивчення дрейфу. Оцінку ймовірності відхилення частоти алеля від вихідного значення можна одержати шляхом використання стандартних відхилень (о). Дисперсія частоти алеля визначається формулою
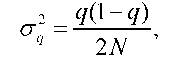
де q - частота алеля а у вихідній (і-тій) генерації, N - чисельність наступної дочірньої (i+1) генерації. Звідки стандартне середньоквадра-тичне відхилення частоти гена в дочірній генерації
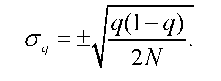
Чим більше відхилення частоти алеля від вихідного значення, тим рідше воно трапляється: до інтервалу від q - о до q + о потрапляє 68,27 % усіх можливих випадкових відхилень частоти алеля, до інтервалу від q - 2о до q + 2о - 95 % і т. д.
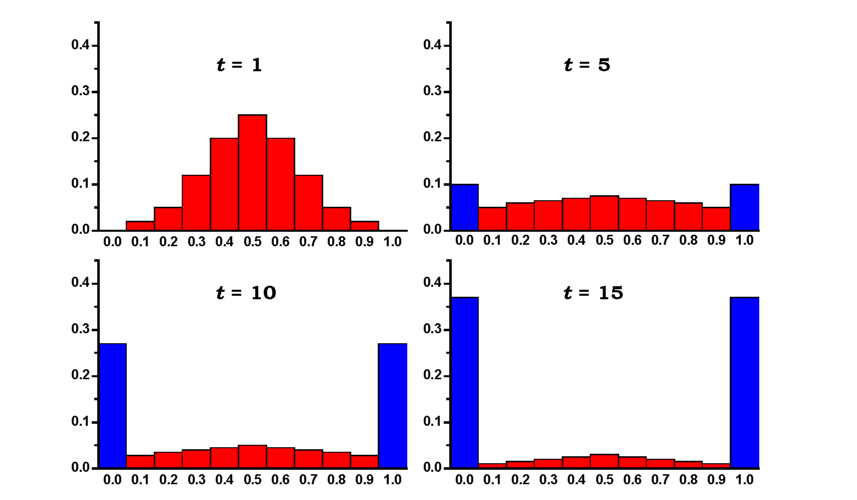
У результаті випадкових змін частот генів під час зміни генерації відбуватимуться як фіксація, так і елімінація алелів, тобто збільшення гомозиготності популяції, втрата мінливості (рис. 8.6, 8.7). Швидкість процесу втрати мінливості дорівнює к = 1/(2N). Інакше кажучи, гетерозиготність у групі з N особин, що розмножуються шляхом випадкових схрещувань, у кожній генерації буде зменшуватися у 2N разів:
 |
|
Рис. 8.6. Криві дрейфу генних частот. По осі абсцис відкладено кількість поколінь, по осі ординат - частоту гена. Криві отримано в комп'ютерних експериментах, де моделювалась популяція з 10 і 100 особин із початковою частотою гена, яка дорівнювала 0,5 |
 |
|
Рис. 8.7. Зміни розподілу генної частоти в результаті дрейфу генів: по осі абсцис - частота алеля, по осі ординат - частка популяцій із відповідною частотою. Популяції складаються з 5 гермафродитних особин; схрещування випадкові: початкова частота гена (при t = 0) дорівнює 0,5. Наведено розподіли для 1, 5, 10 і 15-ї генерацій. Сині стовпці відповідають частці популяцій, у яких алель втрачено або він фіксований (за Кімура, 1985) |
|
Гетерозиготність у групі з N особин після t панміктичних генерацій можна визначити за формулою |
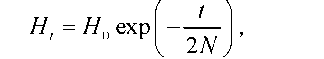 |
де Н - вихідна гетерозиготність, Н - гетерозиготність після t генерацій.
Таким самим чином (тобто зі швидкістю 1/(2N) за генерацію) при генетичному дрейфі змінюється ще один показник мінливості - кількість локусів, за якими не відбулась фіксація або елімінація одного з алелів.
Основними наслідками дрейфу генів є:
• зміна частот алелів у непередбачуваному напрямку;
• збільшення частки гомозигот;
• збіднення генофонду (зникнення алелів).
Дрейф суттєво впливає на генетичну структуру популяції при коливаннях чисельності, коли чисельність популяції різко зменшується (ефект шийки пляшки), а генофонд наступних поколінь визначає невелика група особин - засновників популяції (ефект засновника). Хоча чисельність популяції може потім значно зрости, гени всіх особин походять від невеликої кількості генів, що випадково були присутніми в засновників популяції. Зокрема, це стосується популяцій людини на певних етапах еволюції Homo sapience (розділ 7). Ефект засновника має місце як при проходженні популяції "через шийку пляшки", так і при заселенні видом нових відокремлених територій (наприклад, островів).
Інбридинг - схрещування між генетично спорідненими особинами -є одним із порушень панміксії. При цьому також (як і при дрейфі) зростає гомозиготність популяції, але (на відміну від дрейфу) частоти алелів залишаються незмінними.
Розрізняють кілька різновидів такого схрещування залежно від рівня генетичної спорідненості шлюбних партнерів. Сам термін "інбридинг" - розведення "в собі" - прийшов із тваринництва, де ним називали схрещування в межах однієї лінії; типу брат х сестра тощо. Найтіснішим інбридингом є самозапліднення.
Для оцінки сили інбридингу використовують так званий коефіцієнт інбридингу F - імовірність того, що в генотипі особини обидва алелі певного гену будуть ідентичні за походженням. Розглянемо родовід зі схрещуванням типу брат х сестра (рис. 8.8).
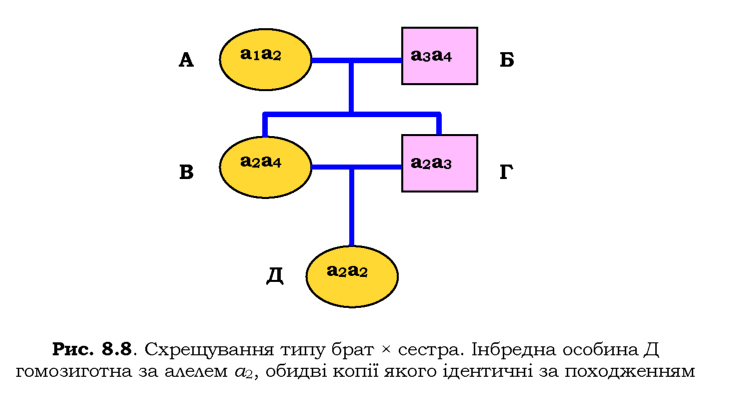
Усі учасники цього родоводу - гомозиготи за рецесивним алелем а, усі копії якого в особин А і Б однакові, але потрапили до них від різних предків і мають різне походження. Позначивши їх для зручності номерами, можна простежити їхню долю в потомстві. Особини В та Г мають різні комплекти алелів, але один із них у них спільний (а2). Саме цей алель міститься в генотипі Д у подвійному екземплярі - це дві копії-репліки одного алеля, що був у генотипі А. Таким чином, Д - гомозигота за алелями, що не тільки однакові, а ще й мають ідентичне походження..
Імовірність того, що А передасть алель а2 обом нащадкам В і Г дорівнює 1/2 х 1/2 = 1/4. Так само ймовірність того, що В і Г одночасно передадуть цей алель нащадку Д становить 1/2 х 1/2 = 1/4. Загальна ймовірність того, що до Д від А потраплять два екземпляри а2, дорівнює (1/2)4 = 1/16. Оскільки в даному родоводі будь-який із чотирьох варіантів алеля а може потрапити до Д у подвійному екземплярі, імовірність того, що Д буде гомозиготою за алелями, ідентичними за походженням, становитиме F = 4 х (1/2)4 = 1/4.
При самозаплідненні F = 1/2: це означає, що частка гетерозигот у кожному наступному поколінні буде вдвічі меншою, ніж у попередньому, а через t поколінь частка гетерозигот становитиме (1/2)(.
У загальному вигляді при інбридингу частка гомозигот буде зростати за рахунок гетерозигот:
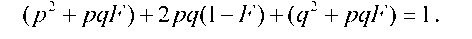 |
|
Значення F може змінюватись від 0 до 1. Якщо в останню формулу підставити F = 0, одержимо рівняння Харді - Вайнберга для пан- |
міктичних популяцій p2 + 2pq + q2 = 1. Значення F = 1 означає повну відсутність гетерозигот у популяції, а частота генотипів АА та аа дорівнюватиме частотам алелів А та а відповідно.
До такого стану популяція може дійти протягом певної кількості поколінь - чим тіснішим є інбридинг, тим швидше. При цьому частоти алелів залишаться незмінними. Отже, основний наслідок інбридингу для структури популяції полягає в тому, що зростає частка гомозигот. Відповідно, зростає ймовірність гомозиготності рецесивних (у тому числі летальних або таких, що викликають захворювання) алелів - відбувається так звана інбредна депресія.
Проте деякі види тварин і рослин, які розмножуються шляхом самозапліднення, є цілком життєздатними. Якщо популяція піддається інбридингу протягом багатьох генерацій, "шкідливі" рецесивні алелі видаляються з неї завдяки добору (про який ітиметься нижче), тобто популяція стає гомозиготною за "корисними" алелями. У такий спосіб інбридинг часто використовують у селекційній роботі з метою отримання рослин і тварин із бажаними ознаками.
Оскільки інбридинг (у разі відсутності добору) не змінює частоти алелів, для оцінки генетичної варіабельності інбредної популяції часто використовують очікувану гетерозиготність - частку особин, які були б гетерозиготами за рівнянням Харді - Вайнберга (2pq для дво-алельного локусу). Так, для самозапильних рослин реальна гетерозиготність може бути дуже низькою, але популяція може характеризуватися досить значною генетичною варіабельністю, мірою якої й буде слугувати очікувана гетерозиготність. Зазвичай очікувану гетерозиготність усереднюють для кількох досліджених локусів.
Мутації є первинним джерелом генетичної мінливості в популяціях. Вони викликають зміни частот як алелів, так і генотипів. Розглянемо випадок, коли внаслідок прямих мутацій алель А перетворюється на а (А ^ а) із частотою и, а зворотні мутації а ^ А відбуваються з частотою v. Тоді за умови певного співвідношення и та v в популяції встановиться рівноважне співвідношення генотипів АА, Аа та аа.
Спочатку оцінимо тиск прямих мутацій. Нехай частота алеля А у вихідному поколінні дорівнює p0. У наступному поколінні ця частота зменшиться за рахунок мутацій:
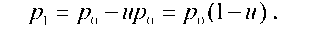
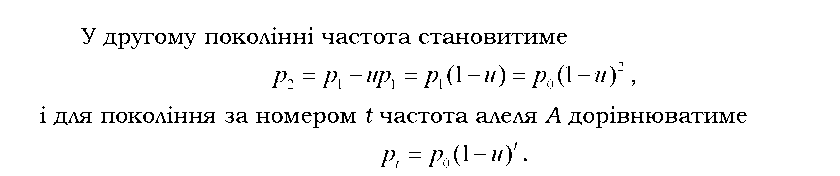
З часом за рахунок мутацій алель А мав би зникнути з популяції. Але цей час досить тривалий: щоб частота алеля змінилась від 1 до 0,99 при u = 10-5 на гамету за покоління, необхідно 1 тис. поколінь. При цьому не враховано вплив зворотних мутацій.
Якщо вихідні частоти алелів А та а становлять відповідно р0 і q0, а частоти прямих і зворотних мутацій - u та v, то в першому поколінні частка алелів А за рахунок прямих мутацій зменшиться, а за рахунок зворотних - зросте:

Міграції, або потік генів між популяціями, приводять до зростання мінливості в межах популяцій і зменшення різниці між ними. При цьому змінюються як частоти алелів, так і частоти генотипів. Масштаби цих змін залежать від розмірів популяцій та інтенсивності міграцій.
Розглянемо популяцію (реципієнт), до якої з певною частотою мігрують особини з інших популяцій (донорів). Вважатимемо, що розмір популяції залишається сталим, іммігранти вільно схрещуються з резидентами й мають однакову життєздатність.
|
Нехай у популяції-реципієнті початкова (доміграційна) частота алеля А становить po, у популяції-донорі - pd. Позначимо частку мігрантів як m. Тоді в генофонді наступного покоління частка генів від мігрантів становитиме m, а від резидентів - (1 - m). Через одне покоління міграції (імміграції) частота алеля в популяції-реципієнті буде |
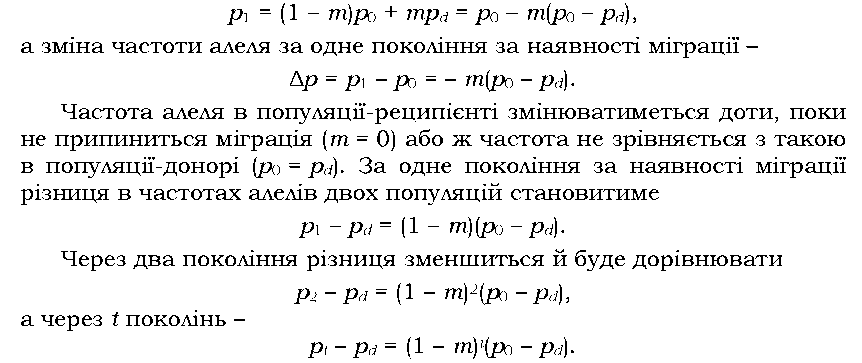 |
Остання формула дає можливість прогнозувати зміну частоти алеля в популяції через певний час після початку міграції, якщо відома інтенсивність цієї міграції m. Слід відмітити, що йшлося саме про імміграцію: еміграція з популяції (у разі відсутності будь-якої генетичної вибірковості, тобто якщо еміграція щодо генотипів буде випадковою, а отже, пропорційною частоті генотипів) до зміни частот алелів не приведе.
Добір можна визначити як диференційне відтворення в популяції особин із різними генотипами. Під впливом добору частоти алелів і частоти генотипів будуть змінюватись у певному напрямку: добір може спричинити втрату чи фіксацію алеля.
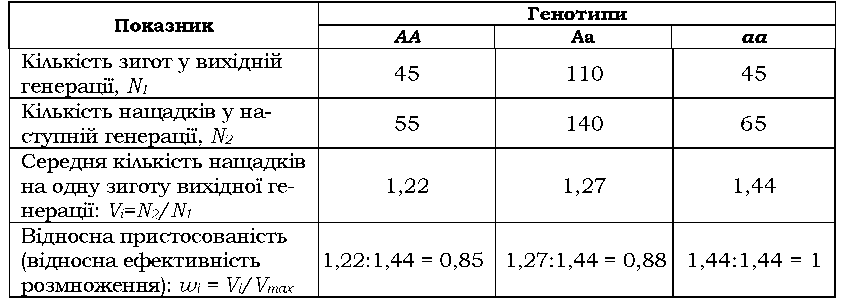
Диференційний внесок особин із різними генотипами у відтворення наступної генерації може бути кількісно виражений через відносну ефективність їхнього розмноження, або відносну пристосованість w, -відношення пристосованості особин із певним генотипом до пристосованості найкращого варіанта, який існує в популяції (табл. 8.3).
З відносною пристосованістю простою залежністю пов'язаний коефіцієнт добору s, який указує, наскільки пристосованість особин даного генотипу є меншою за пристосованість найкращого варіанта:
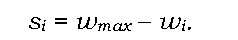
Зрозуміло, що відносна пристосованість найкращого варіанта дорівнюватиме одиниці, а коефіцієнт його добору - нулю.
|
Таблиця 8.3. Приклад визначення відносної пристосованості w трьох генотипів |
 |
Унаслідок того, що особини з різними генотипами матимуть різну пристосованість, протягом ряду генерацій відбуватимуться зміни у співвідношенні генотипів і частот генів до повної елімінації одного з алелів або до такого співвідношення частот генів, при якому в популяції встановиться рівновага. Зміну частоти алеля протягом однієї генерації під впливом добору в загальному випадку показано в табл. 8.4.
|
Таблиця 8.4. Зміна частоти гена під впливом добору протягом однієї генерації |
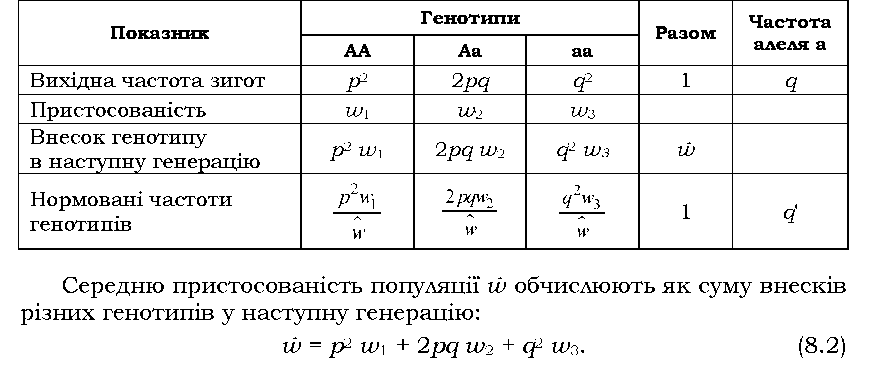 |
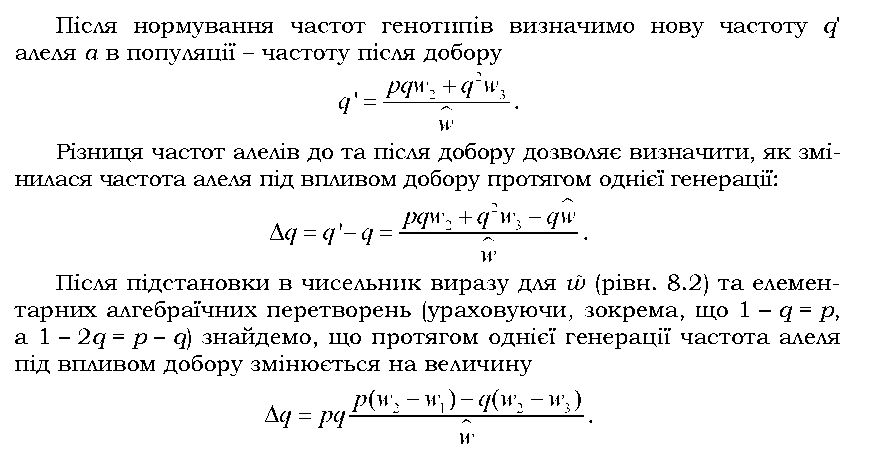
Знак при Aq, який характеризує напрям добору, залежить від співвідношення пристосованості різних генотипів W1, и>2 і и>3 (оскільки р, q і знаменник дробу - завжди додатні числа), а на швидкість добору, інакше кажучи, на величину Aq матимуть вплив як значення пристосо-ваностей, так і значення вихідних частот генів. При Aq = 0 має місце генетична рівновага:
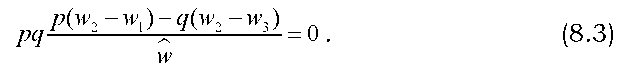
|
Це рівняння рівноваги має два тривіальні розв'язки (p = 0 або q = 0), а також третій розв'язок, який відповідає умові p(w2 - W1) = q(w2 - W3). Звідси маємо співвідношення частот алелів, що відповідає рівноважному стану: |
 |
вання) і Ш1 > W2 > W3 (неповне домінування - пристосованість гетерозиготи є проміжною між пристосованостями гомозигот).
2. р = 0, q = 1 - під впливом добору алель а фіксується (добір на користь алеля а) при W1 = W2 < W3 або W1 < W2 < W3.
„ p w2 - w3 ...
3. — = —2-3 - добір приводить до генетичної рівноваги за рів-
q W2- Wi
нянням 8.4. У випадку наддомінування (W1 < W2 > W3) жоден із алелів не видаляється, оскільки обидва мають перевагу в гетерозиготному стані. Початковий напрямок добору визначається при цьому відносними пристосованостями двох гомозигот, після чого реалізується стан стійкої рівноваги, коли частоти алелів залишаються незмінними. Якщо гетерозигота характеризується пригніченою пристосованістю (недодомінування, W1 > W2 < W3), реалізується нестійка рівновага: частоти алелів залишаються незмінними, поки не здійснюється пертурбація рівноваги за рахунок тих чи інших еволюційних факторів - у цьому випадку рівновага порушується й один із алелів урешті-решт фіксується.
Аналогічно зміні частоти гена при доборі змінюватиметься середня пристосованість популяції w. Величина Aw - зміна середньої пристосованості популяції внаслідок добору протягом однієї генерації -завжди додатна. Отже, у результаті добору середня пристосованість популяції не може зменшуватися - вона може бути постійною або збільшуватися. Відповідно до фундаментальної теореми природного добору Фішера (Ronald Aylmer Fisher), швидкість її збільшення буде тим вища, чим нижче її вихідне значення й чим вища різниця пристосованості між особинами з різними генотипами. У рівноважних точках (Aq = 0) значення Aw дорівнюватиме нулю. За умови стійкої рівноваги в даній точці середня пристосованість популяції буде максимальною.
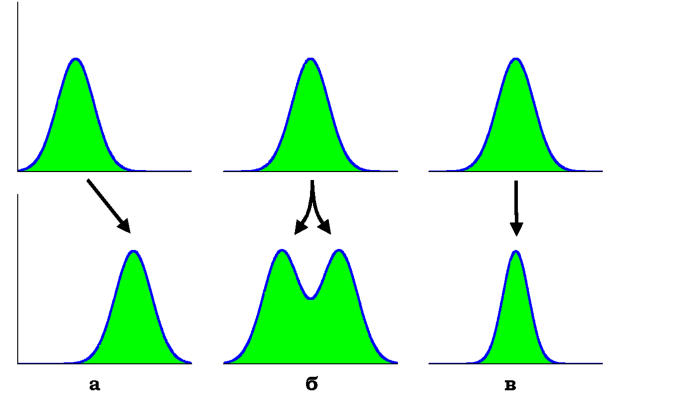
Більшість ознак, що перебувають під дією добору, є кількісними -залежать від кількох генних локусів (див. розділ 3). Добір може змінювати розподіл особин у популяції за значенням кількісної ознаки трьома різними шляхами. Якщо добір сприяє значенням ознаки на одній із меж розподілу, відбувається поступовий зсув розподілу у відповідному напрямі - спрямований добір (рис. 8.9, а). У природі така ситуація реалізується, коли умови середовища змінюються в певного напрямі, примушуючи популяцію адаптуватися до цих змін.
Якщо добір сприяє значенням на обох межах розподілу, відбувається дизруптивний добір: практично, це спрямований добір, що діє в обох напрямках (рис. 8.9, б). Зазвичай дизруптивний добір реалізується або за умов шлюбної асортативності (схрещування переважно відбуваються між особинами з подібними екстремальними проявами ознаки), або внаслідок екологічної чи географічної ізоляції субпопуляцій.
Добір може також сприяти консервації середніх значень розподілу, "відсікаючи" екстремальні варіанти, - стабілізуючий добір
(рис. 8.9, в). Такий процес відбувається, коли особини із середніми проявами ознаки характеризуються підвищеною пристосованістю.
 |
|
Рис. 8.9. Зміна розподілів особин за кількісною ознакою (по осі ординат - частота) під дією добору трьох типів: спрямованого (а), дизруптивного (б), стабілізуючого (в) |
Результуюча динаміка структури популяції залежить від питомого впливу кожного з розглянутих вище факторів, а також від зміни чисельності самої популяції, від змін, які відбуваються в сусідніх популяціях, від зміни умов середовища тощо. У тих випадках, коли зміни частот генотипів і алелів мають довготривалий або навіть незворот-ний характер (як, скажімо, втрата алеля внаслідок дрейфу), має місце елементарна еволюційна подія.
Мутації, ізоляція або міграції, добір, дрейф і асортативність схрещувань лежать в основі мікроеволюції - необоротних змін генетичної структури популяцій. Власне, еволюція і відбувається через процеси мікроеволюції, які здійснюються в межах популяцій і можуть спричинити появу нових видів.
Зміна генетичної структури популяції залежить від складної взаємодії між різними еволюційними факторами (факторами динаміки популяції). Головна причина генетичної гетерогенності популяцій -мутаційний процес, який постачає "вихідний матеріал" для еволюції. Більшість точкових мутацій нейтральні або майже нейтральні: вони або не торкаються кодуючих послідовностей, або приводять до синонімічних замін кодонів, або до таких замін амінокислот, що не мають суттєвих наслідків для структури та функціонування білка. Такі нейтральні мутації випадково фіксуються генетичним дрейфом, що зумовлює зростання генетичної гетерогенності. За постійної швидкості накопичення нейтральних нуклеотидних (чи амінокислотних) замін аналіз варіантів послідовностей у різних таксономічних груп є потужним методом оцінки еволюційних відстаней і філогенетичних зв'язків.
Швидкість результуючого накопичення замін не однакова для різних ділянок геному та різних білків: деякі білки або окремі їхні ділянки (наприклад, такі, що беруть участь у формуванні активних центрів ферментів) відрізняються підвищеною консервативністю. У цьому випадку мутації, які приводять до замін у консервативних ділянках, швидко елімінуються добором. Теорія нейтральної еволюції, сформульована Кімурою та Ота (Motoo Kimura, Tomoko Ohta), відводить головну роль в еволюції випадковим процесам (мутації та дрейф), залишаючи за добором функцію позбавлення від шкідливим мутацій: популяції не "поліпшуються" внаслідок фіксації корисних мутацій, вони просто не стають гіршими, фіксуючи тільки нейтральні або майже нейтральні мутації.
Популяція, що перебуває під тиском еволюційних факторів, які спричиняють генетичну гетерогенність, може раптово розділитися на субпопуляції. Якщо вони виявляються не здатними до схрещувань між собою, то стають репродуктивно ізольованими. Така ізоляція -ключова подія у формуванні нових видів.
Вид можна визначити як групу популяцій, представники яких схрещуються або потенційно здатні схрещуватись між собою, і які репродуктивно ізольовані від інших таких груп. До репродуктивної ізоляції можуть привести різноманітні механізми, їх зазвичай поділяють на дві групи: презиготні (екологічна ізоляція, коли популяції займають різні місця проживання в межах спільного регіону; часова ізоляція - парування або цвітіння відбувається в різні пори року; по-ведінкова ізоляція - відсутній взаємний потяг між статями різних популяцій; гаметна ізоляція - гамети є несумісними тощо) і постзиготні (гібриди не є життєздатними або не здатні давати потомство).
Очевидним шляхом розділення на субпопуляції є, наприклад, географічне розділення: дві популяції на різних островах, вершинах гір, у різних лісових масивах після міграцій тощо. За досить довгий час ізоляції в різних популяціях накопичуються генетичні відмінності, що приводять урешті-решт до репродуктивної ізоляції за одним із пре-або постзиготних механізмів. Навіть якщо дві такі популяції знов займають спільний ареал унаслідок змін навколишнього середовища, схрещування між ними вже стає неможливим: два нові види використовують середовище різними шляхами, добір проти гібридних форм сприяє підсиленню ізоляції та подальшій диференціації.
Контрольні запитання і завдання
1. Дайте визначення популяції. Що таке менделівська популяція? Що таке генетична рівновага?
2. Від чого залежить верхня межа чисельності популяції?
3. Що визначає ефективну чисельність популяції?
4. Як в популяціях оцінюють рівень генетичної мінливості кількісних і альтернативних ознак?
5. Дайте визначення поліморфності та гетерозиготності.
6. Які системи схрещувань ви знаєте?
7. Сформулюйте закон Харді - Вайнберга. За яких умов він виконується?
8. Від чого залежить інтенсивність генетичного дрейфу?
9. Які наслідки дрейфу та інбридингу? У чому їхня відмінність?
10. Що таке коефіцієнт інбридингу? Як його визначити?
11. Що таке "ефект засновника" та "ефект шийки пляшки"?
12. Як мутаційний процес впливає на генетичну структуру популяції? Наскільки суттєвим є вплив мутацій різних типів на генетичну структуру популяцій?
13. Охарактеризуйте особливості впливу міграцій на генетичну структуру популяції? Як частота алеля в популяції залежить від еміграції та імміграції?
14. Як можна оцінити відносну пристосованість генотипу?
15. Що таке коефіцієнт добору?
16. Які типи добору вам відомі? У чому полягають їхні особливості?
17. Що таке мікроеволюція?
18. У чому полягає теорія нейтральної еволюції?
Автор: admin от 18-07-2013, 17:04, Переглядів: 24949