Формальна генетика: закономірності спадкування ознак
Виходячи з розуміння молекулярних і цитологічних основ функціонування апарату спадковості, викладених у попередніх розділах, можна описати процеси передачі кінцевого результату активності генів - ознак - від батьків до нащадків і далі в ряду поколінь. При цьому використовують певну систему позначень, яка дозволяє, відволікаючись від конкретних молекулярних механізмів, записати процес у символьній формі - на кшталт того, як за допомогою математичних формул і рівнянь описують фізичні процеси. Так само, як математична фізика тісно пов'язана з фізикою експериментальною, така формальна генетика є засобом або передбачити характер розподілу ознак у нащадків на основі відомостей про набір генів (генотип) батьків, або навпаки - установити генотип батьків і нащадків за кількісним розподілом ознак.
Історично генетика як наука розпочалася саме з вирішення питань другого типу, і досить довгий час носила характер формальної дисципліни просто тому, що молекулярна природа гена була невідомою. Проте це не завадило встановити важливі генетичні закономірності вже в цей початковий період.
Основні закони спадковості були відкриті Грєгором Менделем (1822-1884), монахом августинського монастиря, який жив у австрійському місті Брюнн (нині Брно, Чехія), викладав математику в монастирській школі та експериментував із передачею у спадок індивідуальних ознак гороху Pisum sativum. Робота Менделя, яка є взірцем наукової роботи, тоді не привернула уваги широкого загалу, оскільки випередила свій час. Менделівські результати були відкриті заново через З5 років, а саме в 1900 р. (див. історичну довідку), відколи й беруть початок систематичні генетичні дослідження.
Одним із головних понять генетики є поняття ознаки - певної визначеної властивості організму або його частини, за якою одна особина відрізняється від іншої. Ознакою в генетичному розумінні можна вважати будь-яку характеристику на рівні зовнішніх рис, фізіологічних або біохімічних особливостей: вага, колір, ріст, набір речовин певного типу в органах і клітинах, сукупність певних білків тощо. Сукупність ознак даного організму називають фенотипом. Мендель усвідомив, що ознаки, які можна використати в генетичному аналізі, повинні виявлятися постійно (незалежно від віку, зовнішніх умов тощо) і мати два або більше альтернативних проявів.
Найважливішим експериментальним підходом генетики (практично єдиним на початковому етапі 'її розвитку) є схрещування - природне або штучне поєднання двох гамет при заплідненні. Схрещування позначають знаком множення "х". У схемах на перше місце ставлять генотип жіночої статі, яку позначать символом "?" (дзеркало Венери), на друге - чоловічої із символом "<^" (щит і спис Марса). Під генотипом у схемах схрещувань розуміють не сукупність усіх генів організму, а тільки тих, що стосуються ознак, які аналізуються.
Домінантні та рецесивні алелі (див. розділ 1) одного гена позначають, як правило, однаковими великою та малою літерами відповідно. Потомство від схрещування двох особин із різними альтернативними проявами ознаки називають гібридним, а окремого представника такого потомства - гібридом. Батьківські організми, які беруть у схрещування, позначають літерою Р (від латинського parentes - батьки), а потомство - літерою F (від латинського filii - діти). Зазвичай біля літери F ставлять цифровий індекс, який відповідає порядковому номеру гібридного покоління.
Схрещування, в якому проводиться аналіз однієї пари альтернативних ознак, називається моногібридним. Повернемося до прикладу, що вже розглядався наприкінці розділу 1 - спадкування забарвлення насіння гороху, що залежить від гена sgr. Відповідно до сформульованих вище правил, позначимо нормальний (домінантний) алель цього гена як А (ген дає функціональний продукт, який зумовлює руйнування хлорофілу та, відповідно, жовтий колір насіння), а мутантний рецесивний алель (ген не дає функціонального продукту, і насіння залишається зеленим) як а. Власне, саме така мутація, коли ген втрачає здатність кодувати повноцінний продукт, і зумовлює зазвичай виникнення рецесивної форми цього гена.
 |
|
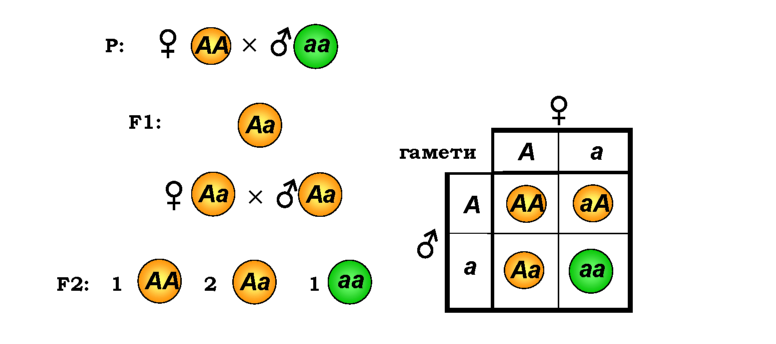
Рис. 3.1. Схема моногібридного схрещування гомозиготних батьківських форм гороху з жовтим і зеленим насінням і наступного схрещування гібридів першого покоління. Праворуч показано решітку Пеннета для другого схрещування |
Мендель узяв для схрещування дві попередньо отримані ним чисті (генетичні) лінії - так називають групи особин, які стабільно відновлюють у низці поколінь одні й ті самі спадково константні ознаки. Тобто батьківські форми (Мендель установив це за кінцевим результатом) були гомозиготами за алелями А та а із насінням відповідно жовтого та зеленого кольору (рис. 3.1). У першому поколінні після об'єднання гамет двох типів (кожен із батьків продукує гамети тільки одного типу) усі особини були гетерозиготами жовтого кольору - виявлявся тільки домінантний алель, а ознака іншого батька (рецесивна)
не спостерігалася. Це явище стало основою для формулювання першого закону Менделя (або закону одноманітності гібридів першого покоління): у першому поколінні від схрещування гомозигот із домінантною та рецесивною ознаками виявляється тільки домінантна ознака.
Гібриди першого покоління продукують уже два типи гамет -з алелями А або а. При схрещуванні цих гібридів мають утворитися три різні комбінації алелів (строго кажучи, чотири комбінації, дві з яких - аА та Аа - є еквівалентними). Для полегшення розрахунків цих комбінацій іноді застосовують таблицю, запропоновану Пеннетом ('її ще називають решіткою Пеннета): у першому рядку та першому стовпчику записують типи гамет, і тоді на перетині отримують комбінації алелів у нащадків. Із решітки Пеннета на рис. 3.1 видно, що частота гетерозигот Аа буде у два рази вищою, ніж однакові частоти гомозигот за домінантним і рецесивним алелями. Отже, для великої кількості особин другого покоління генотипи АА, Аа, аа будуть отримані у співвідношенні 1 : 2 : 1, а співвідношення фенотипів (жовтих до зелених) буде дорівнювати 3 : 1. Саме цей результат і отримав Мендель. Твердження про те, що у другому поколінні відбувається розщеплення домінантної ознаки на домінантну та рецесивну у співвідношенні 3 : 1, називають другим законом Менделя, або законом розщеплення.
Отримані групи нащадків Мендель піддав самозапиленню та встановив, що рослини з рецесивними ознаками не дають розщеплення в наступних поколіннях. Таким самим чином поводить себе й 1/3 всіх особин із домінантною ознакою. А решта 2/3 домінантних особин дають розщеплення на домінантних і рецесивних знову ж таки у співвідношенні 3 : 1.
Не будемо тут ще раз повторювати висновки, які зробив Мендель на підставі отриманих ним результатів, - практично, він описав поведінку хромосом під час мейозу та запліднення. Важливість його роботи полягала ще й у демонстрації того, що, слідкуючи за спадкуванням зовнішніх ознак при схрещуваннях, можна робити висновки щодо генотипу особини.
Іноді такі висновки можна робити й без схрещувань: у разі наявності в певного організму рецесивної ознаки він завжди є гомозиготою. Але у випадку домінантної ознаки такий однозначний висновок зробити неможливо: особина може бути як гомо-, так і гетерозиготою. У схемах схрещувань таку особину часто записують у вигляді фенотипового радикала А_ (маючи на увазі, що замість _ може стояти як велика, так і маленька літера - без зміни при цьому фенотипового прояву). Для встановлення генотипу застосовують схрещування досліджуваної особини (з домінантною ознакою) з особиною, гомозиготною за рецесивом, -аналізуюче схрещування, яке чітко ідентифікує генотип особини. Залежно від генотипу досліджуваної особини в аналізуючому схрещуванні або спостерігається розщеплення 1 : 1 (гетерозигота, варіант 1), або всі нащадки характеризуються домінантною ознакою (гомозигота, варіант 2):
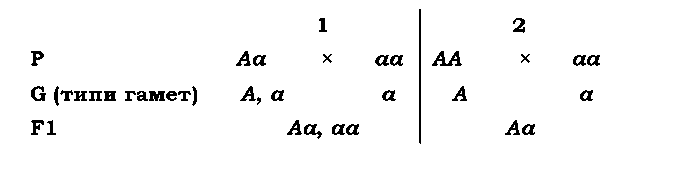
Описаний приклад спадкування забарвлення насіння гороху належить до ситуації так званого повного домінування: прояв ознаки не залежить від кількості (два або один) домінантних алелів у генотипі. Проте у природі часто зустрічаються ще інші типи взаємодії алельних генів.
Перший із них спостерігається у випадку, коли рецесивний алель не дає функціонального продукту, але ознака залежить від кількості копій домінантного алеля в генотипі. Наприклад, забарвлення квітки в червоний колір у ротиків Antirrhinum majus залежить від кількості червоного пігменту, а цей останній з'являється внаслідок активності ферменту, що контролюється певним геном. Два гени АА зумовлюють інтенсивне червоне забарвлення, гомозигота аа (відсутність ферменту та, відповідно, пігменту) характеризується білим кольором квітки, гетерозигота має вдвічі меншу інтенсивність забарвлення порівняно з гомозиготою за домінантним алелем (удвічі менше ферменту й пігменту) -квітку рожевого кольору (рис. 3.2). Аналогічно спадкується кучерявість пір'я в голубів: особина, гетерозиготна за геном кучерявості, характеризується хвилястим пір'ям. Такий тип взаємодії алельних генів називають неповним домінуванням. За неповного домінування гетерозигота характеризується фенотипом, відмінним від обох гомозигот -із цією поправкою перший закон Менделя залишається без змін. Що стосується результатів схрещування гетерозигот, то розщеплення за генотипом і фенотипом збігатимуться та становитимуть 1 : 2 : 1.

Аналогічні розщеплення характерні й для кодомінування з тією різницею, що нащадки від схрещування гомозигот різних типів будуть характеризуватися наявністю фенотипових ознак обох батьків одночасно. Тобто в цьому разі обидва алелі є домінантними: дають функціональні продукти, що трохи розрізняються за певними характеристиками. Така ситуація досить часто спостерігається для ферментів, коли два алелі зумовлюють синтез двох білків - ізоферментів -із практично однаковою активністю, але при цьому вони розрізняються за молекулярною вагою (один із білків має додатковий структурний домен, кінцевий хвіст тощо).
Розглянуті приклади потребують важливого уточнення. З того факту, що у двох гомологічних хромосомах можуть міститися лише два різні алелі одного гена, не випливає, що у групі особин ген обов'язково має максимум два алелі. Насправді, у більшості випадків гени існують у вигляді кількох (теоретично необмеженої кількості) різних алелів. Це явище отримало назву множинний алелізм. Одним із прикладів множинного алелізму є система груп крові АВ0. Добре відомі чотири групи крові - 0, А, В і АВ - визначаються трьома алелями одного гена: ІА, ІВ, і0. Алелі ІА та ІВ є кодомінантними (відповідають за формування на поверхні еритроцитів антигенів двох типів - А і В), але при цьому домінують над алелем і0, який не продукує жодного ан-тигену. За наявності трьох алелів можливо шість комбінацій між ними, але рецесивність Р зводить кількість фенотипів (груп крові) до чотирьох: групі 0 відповідає генотип і0і0, групі А - генотипи ІАІА та ІАі0, групі В - ІВІВ та 2е і0, групі АВ - генотип ІАІВ.
Дигібридні й полігібридні схрещування
У своїх дослідах Мендель також аналізував спадкування двох пар альтернативних ознак одночасно: такі схрещування називаються ди-гібридними, а особини, що є гетерозиготними за двома генами одночасно, - дигетерозиготами. Мендель схрещував форми гороху з жовтим і гладеньким насінням із рослинами з насінням зеленим та зморшкуватим. Усі гібриди першого покоління мали жовті й гладкі горошини: жовтий колір, як і в попередньому прикладі, домінував над зеленим, а гладка форма - над зморшкуватою. Залишивши позначення А/ а за алелями забарвлення, алелі форми насіння позначимо як B (гладенька) і b (зморшкувата). Після самозапилення гібридів першого покоління спостерігалося розщеплення 9 : 3 : 3 : 1 - виявилося, що 9/16 потомства характеризувалось наявністю двох домінантних ознак одночасно (жовте та гладке насіння), дві групи по 3/16 - наявністю лише однієї з двох домінантних ознак (жовте - зморшкувате, зелене - гладке), і 1/16 особин були притаманні обидві рецесивні ознаки.
Якщо припустити (що й зробив Мендель), що два гени спадкують-ся незалежно, то дигібридні схрещування можна розглядати як два незалежні моногібридні схрещування. За другим законом Менделя, розщеплення за кольором у другому поколінні становить 3/4А_: 1/4аа, і за формою також - 3/4В_ : 1/4bb. За відомим правилом добутку ймовірностей (імовірність здійснення двох подій одночасно дорівнює добутку ймовірностей кожної з цих подій) можна розрахувати вищезгадане розщеплення:
Використовуючи правило добутку та знаючи розщеплення при моно-гібридному схрещуванні, можна розрахувати співвідношення як феноти-пових, так і генотипових класів у полігібридних схрещуваннях (табл. 3.1).
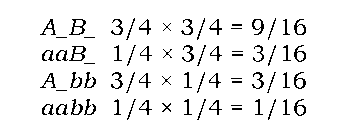
|
Схрещування |
Кількість пар алелів, за якими різняться батьки |
Кількість типів гамет, які може продукувати гетерозигота |
Кількість фенотип-них класів F2 при повному домінуванні |
Кількість генотипних класів F2 |
Частка особин F2, гомозиготних по всіх генах |
|
Моногібридне |
1 |
2 |
2 |
3 |
1/4 |
|
Дигібридне |
2 |
4 |
4 |
9 |
1/16 |
|
Тригібридне |
3 |
8 |
8 |
27 |
1/64 |
|
Полігібридне |
n |
2n |
2n |
3n |
1/4n |
Проаналізувавши результати дигібридних схрещувань, Мендель сформулював постулат про незалежне спадкування ознак, який потім став називатися третім законом Менделя.
ВІДХИЛЕННЯ ВІД МЕНДЕЛІВСЬКИХ РОЗЩЕПЛЕНЬ
Закони Менделя мають для генетики приблизно таке значення, що й закони поведінки ідеальних газів - для термодинаміки. Подібно до того, як модель ідеального газу ускладнюється з метою врахування особливостей реального газу, менделівські закономірності є базисом для розробки складніших моделей, в яких беруться до уваги різноманітні особливості функціонування спадкового апарату в конкретних випадках. Описані вище розщеплення, які реально спостерігаються для певних ознак гороху, є скоріше винятком із правила. Власне, уся постменделівська генетика присвячена з'ясуванню причин різноманітних відхилень від законів Менделя.
Установлення факту відхилення: критерій х2
Перш ніж з'ясовувати причини та механізми відхилень, у кожному конкретному випадку слід відповісти на питання, чи дійсно має місце таке відхилення від очікуваного за Менделем розподілу ознак. Адже відсутність відповідності між очікуваним та реальним співвідношенням фенотипів може бути викликана просто випадковими відхиленнями. Проілюструємо це на простому прикладі підкидання монеток з наступним падінням орлом або решкою. Зрозуміло, що ймовірність падіння на орла / решку становить 1/2, тобто очікуване співвідношення падінь дорівнює 1 : 1. Конкретно на 10 підкидань має бути приблизно 5 падінь на орла та 5 - на решку. Ключовим у попередньому реченні є слово "приблизно" - чим більше буде підкидань, тим ближчим до очікуваного буде результат. Через випадкові відхилення зазвичай (у чому можна легко переконатися) спостерігаються співвідношення на кшталт З-7 падінь на орла та 7-З на решку (а 7 : З = 2,ЗЗЗ:1, що суттєво відрізняється від теоретично очікуваного співвідношення).
Отже, випадкові відхилення можуть приводити до різниці між теоретично очікуваними розщепленнями та такими, що спостерігаються в досліді. Чим більший розмір вибірки, тим меншими будуть такі відхилення. Якщо експериментальні дані відрізняються від теоретично очікуваних тільки за рахунок випадкових відхилень, то кажуть, що виправдовується нульова гіпотеза. У протилежному разі необхідно запропонувати інше пояснення розщепленням, які спостерігаються, а отже, запропонувати іншу гіпотезу.
Для оцінки нульової гіпотези в математичній статистиці користуються х2-критерієм. Величина х2 кількісно відображає відхилення від очікуваного розподілу з урахуванням розміру вибірки:
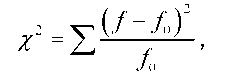
де f - кількість особин з фенотипами певного класу у вибірці, f - очікувана кількість, знак суми вказує на підсумовування по всіх фено-типових класах (числом n). Припустимо, наприклад, що у схрещуванні гороху з жовтим і зеленим насінням (рис. З.1) у другому поколінні отримано 70 рослин з жовтим насінням і ЗО - із зеленим (n = 2). Нульова гіпотеза передбачає співвідношення З : 1, тобто очікувані кількості становлять відповідно 75 і 25. Застосування наведеної формули дає
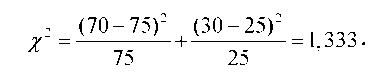
Важливою характеристикою є також число ступенів свободи. Серед усіх фенотипових класів імовірність появи одного з них однозначно визначається ймовірностями решти класів, тобто число ступенів свободи m = n - 1. У нашому прикладі кількість фенотипових класів n = 2, а число ступенів свободи m = 1: для певної вибірки кількість особин одного класу автоматично дає кількість особин іншого класу. Число ступенів свободи важливо враховувати, оскільки зі збільшенням цієї величини росте й імовірність випадкового відхилення від очікуваних величин.
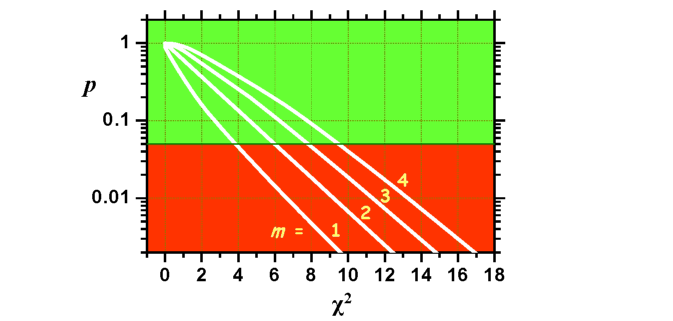
Після визначення числа ступенів свободи необхідно інтерпретувати значення х2 стосовно ймовірності (1 - р) того, що відхилення від очікуваного розподілу є статистично значущими (не випадковими), - насправді, саме таке завдання найчастіше стоїть перед дослідником. Величина р залежить від значення х2 і числа ступенів свободи - зазвичай р установлюють, використовуючи спеціальні таблиці або графіки. На рис. 3.3 наведено у графічній формі табличні значення залежності р від х2 для різних значень числа ступенів свободи. Якщо, наприклад, провести вертикальну лінію з точки 1,333 на осі абсцис до перетину з графіком для m = 1, то можна оцінити, що в нашому прикладі р ~ 0,27.
 |
|
Рис. 3.3. Залежність імовірності р від х2 для чотирьох значень числа ступенів свободи m. Зеленим зафарбовано зону p > 0,05, червоним - p < 0,05 |
Отриману величину р порівнюють з певним рівнем значущості а, який задає прийнятну для даного випадку ймовірність хибного визнання нульової гіпотези невірною: якщо р > а, нульову гіпотезу приймають; якщо р < а, - відкидають, визнаючи відхилення, що спостерігаються, статистично значущими. Зазвичай у біологічних дослідженнях використовують а = 0,05. Тобто, якщо р < 0,05, то з імовірністю > 0,95 (так звана довірча ймовірність 1 - а) відхилення, що спостерігаються, визнаються статистично значущими; для всіх інших значень р береться нульова гіпотеза. Отже, у нашому прикладі нульова гіпотеза виправдовується. У попередньому простому прикладі з підкиданням монети (також одна степінь вільності ) при семи випадіннях орла та трьох - решки х2 = 1,6 і р ~ 0,2 - нульова гіпотеза (у справедливості якої в даному випадку ніхто не сумнівається) також виправдовується.
|
Таблиця 3.2. Величини х2 для двох значень р залежно від числа ступенів свободи |
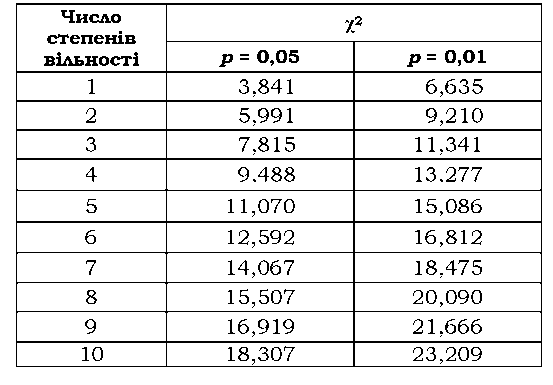 |
На практиці з метою перевірки нульової гіпотези порівнюють розраховане значення х2 із таким критичним значенням, що відповідає р = а (рис. 3.3, табл. 3.2), і приймають нульову гіпотезу, якщо х2 є меншим за критичний, та відкидають у протилежному випадку.
Причини статистично значущих відхилень від менделівських розщеплень
Слід зауважити, що, як це зрозуміло з описаних у розділі 1 цитологічних основ спадковості, менделівські розщеплення у принципі можуть бути реалізованими тільки для видів із нормальним статевим процесом, диплоїдністю обох статей та за умови відсутності порушень у мейозі.
Але й при виконанні зазначених умов випадки відхилень від менде-лівських розщеплень досить численні. При цьому менделівські закономірності не порушуються - просто на них накладаються інші ефекти. Три такі ефекти - взаємодія неалельних генів, зчеплення груп генів в одній хромосомі та розташування гена в статевих хромосомах - розглядаються в окремих підрозділах. Серед інших причин слід назвати такі:
1. Летальність певних комбінацій алелів. Наприклад, при схрещуванні жовтих (гетерозиготних) мишей, розщеплення на жовті й чорні (рецесивні гомозиготи) завжди становить 2 : 1. Подібний результат спостерігається також при схрещуванні платинових лисиць. Якщо перевірити вагітних самок, то виявляється, що чверть ембріонів гине ще до народження. А саме, гинуть гомозиготи за домінант-ним алелем, чому й спостерігається відхилення від очікуваного розщеплення 3 : 1 (відсутній один із чотирьох генотипових класів). Таким чином, у цьому прикладі домінантний алель є насправді домінантним лише щодо забарвлення шерсті - стосовно життєздатності той самий алель є рецесивним. Наведений випадок, коли один ген впливає на декілька ознак одночасно (забарвлення шерсті та життєздатність), є одним із прикладів плейотропії.
2. Існують алелі, які фенотипово виявляються тільки в певної частини організмів, що містять їх у генотипі. Для характеристики такого варіабельного прояву генів використовують поняття пенетрантності та експресивності. Мірою пенетрантності є частка особин, які характеризуються певним фенотипом, серед усіх особин з однаковим генотипом. Деякі спадкові хвороби людини, наприклад подагра, характеризуються неповною пенетрантністю, тобто не всі особи, які несуть мутацію, хворіють. Експресивність відображає силу прояву мутантного гена у фенотипі. Так, дрозофіли, гомозиготні за мутантним алелем eyeless, можуть мати різну кількість фасеток очей - від нормальної кількості до повної їхньої відсутності. Здатність генотипу виявлятися по-різному залежно від зовнішніх умов відображає його норму реакції - певний діапазон прояву, у межах якого ознаки можуть варіювати у відповідь на варіації умов розвитку організму.
3. Деякі ознаки притаманні особинам протягом не всього життя, а виникають лише в певний період: характер розщеплення може залежати від віку. Наприклад, у людини є спадкові синдроми, які виявляються лише в похилому віці (синдром Альцгеймера, хвороба Гентінгтона).
4. До відхилень від очікуваних менделівських розщеплень приводить також явище імпринтингу. Основою його є так звана епігенетична спадковість - успадкування від батьків не просто ДНК, а хроматину, який несе на собі певні хімічні маркери, від яких залежить структурний стан ділянки та, відповідно, активність певних генів (див. розділ 6). Унаслідок такого ефекту прояв певного гена може бути зумовлений тим, від кого саме з батьків дана хромосома отримана.
5. У природі існує багато видів зі статевим диморфізмом, і стать відіграє велику роль у прояві цілої низки ознак (див. підрозділ, присвячений зчепленню зі статтю, а також розділ 6).
Насправді, усі зазначені вище ефекти можуть бути зведені до взаємодії генів у складній системі функціонування геному, хоча й не завжди таку взаємодію можна легко описати: часто треба враховувати надто багато окремих елементів, які впливають один на одного.
Під взаємодією неалельних генів у вузькому значенні зазвичай розуміють випадки, коли декілька різних генів впливають на розвиток однієї ознаки. При цьому також спостерігаються відхилення від очікуваних співвідношень фенотипових класів. Слід розуміти, що йдеться не про безпосередню взаємодію між генами: прояв ознаки залежить від продуктів різних генів (наприклад, ферментів, які залучені до одного біохімічного каскаду). Розрізняють три типи взаємодії неалельних генів: комплементарність, епістаз і полімерію.
Комплементарність - тип взаємодії неалельних генів, при якому присутність у зиготі домінантних алелів цих генів (хоча б по одному для кожного гена) зумовлює такий прояв ознаки, який не спостерігається за наявності лише одного з домінантних алелів окремо. Гени, що взаємодіють за такою схемою, називають комплементарними. Конкретний вид розщеплень на фенотипові класи за комплементарної взаємодії залежить від того, чи мають домінантні алелі кожного з генів власний фенотиповий прояв.
Розглянемо найпростіший випадок взаємодії двох комплементарних генів A/ a і В/ b. Згідно з третім законом Менделя, при дигібрид-ному схрещуванні очікуваним розщепленням за фенотипом у F2 є 9А_В_ : 3А_ЬЬ : 3aaB_ : 1aabb. Саме це розщеплення і спостерігається при спадкуванні форми гребня в курей (рис. 3.4), коли кожен із домінантних алелів двох комплементарних генів має свій власний фенотиповий прояв (табл. 3.3).


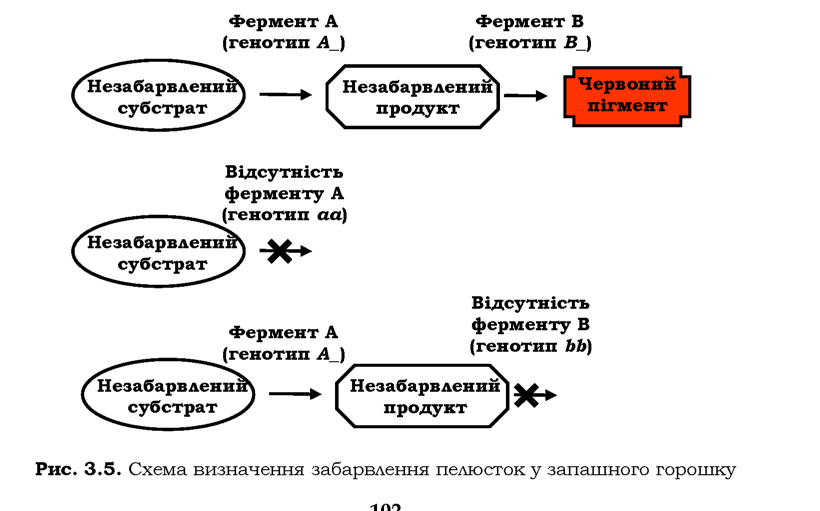
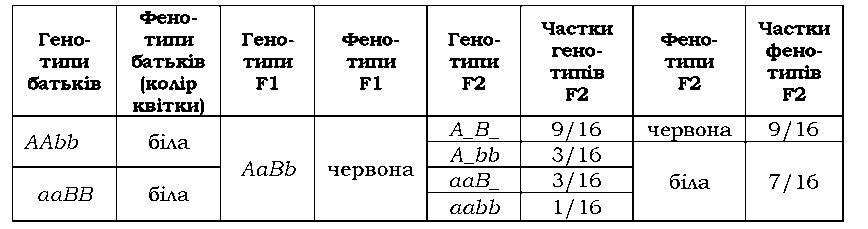
У запашного горошку забарвлення квітки в червоний колір залежить від двох домінантних алелів, жоден із яких не має свого власного фенотипового прояву. Гени кодують два ферменти, залучені до послідовного двоступеневого перетворення певного субстрату на червоний пігмент: у відсутності хоча б одного з ферментів (хоча б одного домінантного алеля) пелюстки квітки залишаються незабарвленими (рис. З.5). Тобто генотипові класи А_ЬЬ, aaB_ і aabb мають однаковий фенотип (білі пелюстки) і формують один фенотиповий клас: у F2 спостерігається розщеплення 9 : 7 (табл. З.4).
|
Таблиця 3.4. Комплементарна взаємодія генів, які визначають забарвлення квітки в запашного горошку |
 |
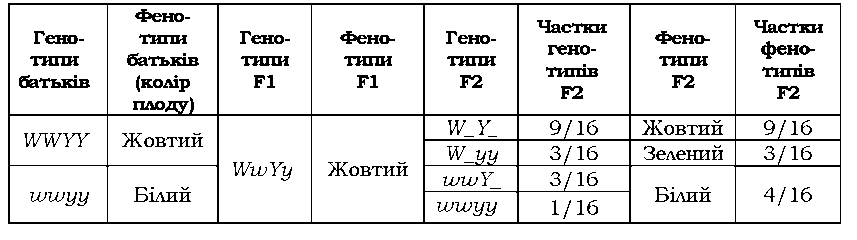
Забарвлення плоду в гарбуза також визначається двома генами (за схемою, аналогічною до такої на рис. 3.5), але тільки один із домінантних алелів цих генів не має власного прояву - у F2 спостерігається розщеплення 9 : 3 : 4 (табл. 3.5). У даному випадку алель W - один із генів, який відповідає за синтез або накопичення зеленого пігменту (хлорофілу). У присутності алеля Y зелений пігмент перетворюється на жовтий (тобто, алель Y у фенотипі виявляється виключно за наявності W, але власного фенотипового прояву не має). У присутності двох алелів w пігмент узагалі не утворюється, тому плоди залишаються білими.
|
Таблиця 3.5. Комплементарна взаємодія генів, які визначають забарвлення плоду в гарбуза |
 |
Ще один тип розщеплення у другому поколінні при комплементарній взаємодій генів спостерігається тоді, коли обидва домінантні алелі мають однаковий власний фенотиповий прояв. Так, домінантні алелі двох певних генів визначають однакову сферичну форму плоду в гарбуза, якщо вони присутні в генотипі окремо; сумісна дія цих алелів зумовлює дископодібну форму. У другому поколінні спостерігаються три фенотипові класи у співвідношенні 9 : 6 : 1 за рахунок об'єднання генотипових класів A_bb і aaB_ (табл. 3.6).

Епістаз - тип взаємодії неалельних генів, при якому один ген здатен пригнічувати дію іншого. Ген, який пригнічує, називають епі-статичним, а ген, дія якого пригнічується, - гіпостатичним. Характер розподілу фенотипових класів залежить від наявності власного прояву у двох генів, що взаємодіють, та від того, домінантний чи рецесивний алель гена є епістатичним.
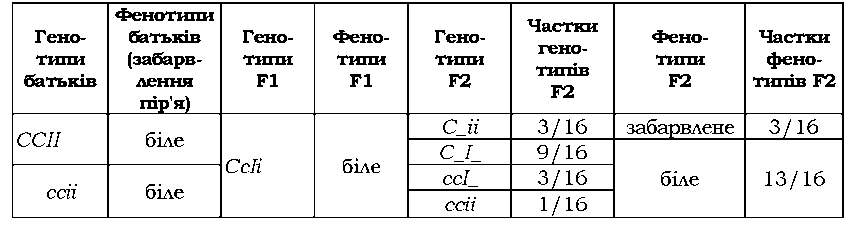
Якщо інгібітором є домінантний алель епістатичного гена, має місце домінантний епістаз (табл. З.7 - алель І пригнічує прояв алеля С при забарвленні пір'я в курей).
|
Таблиця 3.7. Епістатична взаємодія генів, які визначають забарвлення пір'я в курей |
 |
Іншим варіантом розщеплення при домінантному епістазі може бути розщеплення 12 : З : 1. Це спостерігається при забарвленні хутра в деяких гризунів. Алель А зумовлює синтез чорного пігменту, а алель В -рудого. За відсутності обох домінантних алелів хутро залишається білим. За наявності двох алелів (А_В_) синтезуються обидва пігменти, але рудий маскується чорним - хутро в цьому разі має чорне забарвлення.
У випадку, коли гіпостатичний ген не виявляється за присутності епістатичного рецесивного алеля в гомозиготному стані, говорять про рецесивний епістаз. Як приклад даного типу взаємодії можна навести спадкування кольору шерсті в лабрадорських мисливських собак. Один ген зумовлює тип пігменту, який синтезується клітинами шкіри: алель B зумовлює синтез чорного пігменту, а рецесивний алель b -коричневого. Інший ген контролює накопичення пігменту у волоссі: наявність домінантного алеля E приводить до накопичення пігменту, тоді як у гомозигот за рецесивниим алелєм e пігменту у волоссі немає, такі собаки мають жовтий колір. Отже, алель e в гомозиготному стані приводить до відсутності пігменту у волоссі, тобто можна сказати, що він пригнічує прояв гена забарвлення. Очевидно, що генотипові класи B_ee та bbee будуть об'єднуватися в один фенотиповий, і при схрещуванні дигетерозигот спостерігатиметься розщеплення 9 (B_E_ - чорні) : 3 (bbE_- коричневі) : 4 (3 B_ee + 1 bbee - жовті). Очевидно також, що наведений приклад рецесивного епістазу скоріше можна інтерпретувати як комплементарність: чорне забарвлення волосся спостерігається тільки за наявності домінантних алелів обох генів (див. аналогічне розщеплення в табл. 3.5).
Поряд із простим рецесивним епістазом, іноді формально розглядають подвійний рецесивний епістаз - ситуацію, коли рецесивний алель будь-якого з генів, що взаємодіють, у гомозиготному стані є епі-статичним щодо іншого гена. Так, розглянуте розщеплення 9 : 7 (спадкування забарвлення квітки в запашного горошку, табл. 3.4) можна інтерпретувати як подвійний рецесивний епістаз: aa пригнічує прояв алеля B, а bb, у свою чергу, - прояв A. Наведені приклади демонструють, що рецесивний епістаз (простий або подвійний) насправді зводиться до комплементарності.
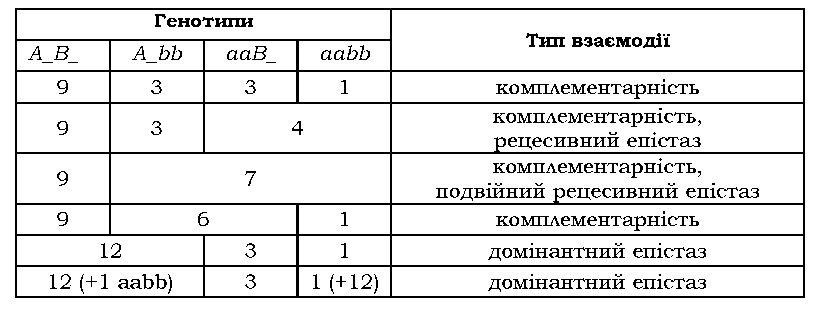
Чітко визначити тип взаємодії неалельних генів можливо в тих випадках, коли відомий біохімічний механізм прояву досліджуваної ознаки (як, скажімо, у випадку забарвлення квітки в запашного горошку). Часто біохімічний механізм залишається невідомим, і тоді обмежуються констатацією формальної генетичної схеми спадкування ознаки. Отже, ідентифікація того чи іншого типу взаємодії неалельних генів часто є умовною. Як показано в табл. 3.3-3.7, при відхиленнях у розщепленнях, зумовлених взаємодією двох генів, співвідношення фенотипових класів, що спостерігаються у F2, завжди можна вивести з класичного 9 : 3 : 3 : 1. Для цього важливо усвідомити, які саме класи об'єднуються, після чого можна зробити висновок про можливий тип взаємодії (табл. 3.8).
|
Таблиця 3.8. Співвідношення фенотипових класів у F2 дигібридного схрещування при деяких типах взаємодії неалельних генів |
 |
Полімерія - тип взаємодії неалельних генів, при якому ознака формується в результаті дії кількох генів з однаковим впливом на ознаку (однозначних генів), тобто йдеться про родини ідентичних або гомологічних генів, що повторюються в геномі та мають однаковий прояв. Сумарний ефект може залежати просто від наявності хоча б одного такого гена (при визначенні якісних ознак) або від кількості домінантних алелів взаємодіючих генів (при визначенні кількісних ознак). Відповідно, розрізняють некумулятивну та кумулятивну полімерію. У разі полімерії прийнято позначати взаємодіючі гени однією буквою, різні гени при цьому позначають нижнім цифровим індексом (А1а1А2а2Аэа3...). При некумулятивній полімерії достатньо одного домінантного алеля будь-якого гена з числа тих, які впливають на ознаку, щоб ця ознака виявилась. Характерне розщеплення у другому поколінні 15 : 1, прикладом є форма плоду у грициків Capsella bursa-pastoris (табл. 3.9).
|
Таблиця 3.9. Полімерна взаємодія генів, які визначають форму плоду в Capsella. bursa-pastoris |
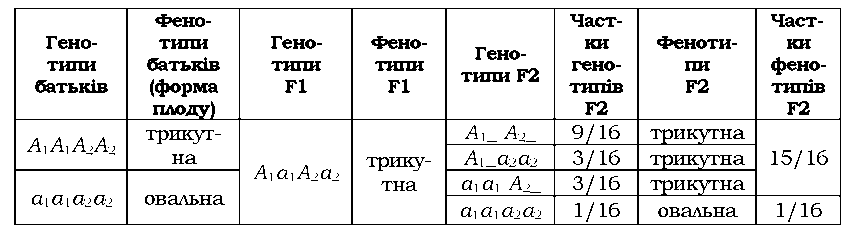 |
У випадку кумулятивної полімерії ступінь прояву ознаки (яку називають адитивною) залежить від кількості домінантних алелів полімерних (адитивних) генів: можна сказати, що вплив окремих генів накопичується, і чим більше домінантних алелів різних генів, тим сильніше проявляється ознака. Саме за таким принципом спадкується, наприклад, колір шкіри в людини (рис. 3.6).

Для двох пар адитивних генів характерне розщеплення у другому поколінні становить 1 : 4 : 6 : 4 : 1 - прикладом є розвиток забарвлення зерна пшениці (табл. 3.10). За типом кумулятивної полімерії спадкується більшість кількісних ознак.
|
Таблиця 3.10. Полімерна взаємодія генів, які визначають забарвлення зерна пшениці |
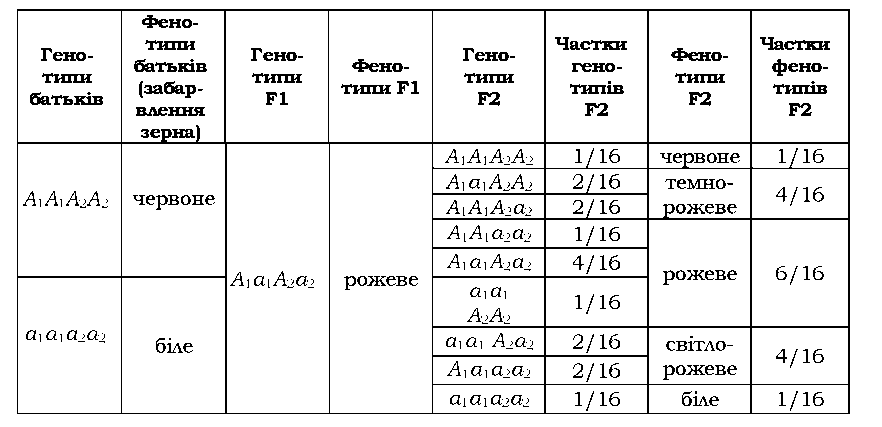 |
Вище йшлося головним чином про ознаки, які чітко виявляються у фенотипі й досить легко відрізняються від альтернативних. При розчепленні їх не виникає сумнівів, до якого фенотипового класу за даною ознакою слід віднести ту чи іншу особину. Такі ознаки (це, наприклад, особливості забарвлення чи форми, відсутність певного ферменту та ін.) зазвичай називають якісними. Однак багато спадкових ознак неможливо точно і якісно описати. Скажімо, коли спостерігаються поступові малопомітні переходи між особинами, а при розщепленні немає чітко розмежованих фенотипових класів. Ці ознаки (вага й розміри тіла, плодовитість, урожайність, продуктивність, скоростиглість, вміст білків і жирів тощо) доводиться вивчати шляхом вимірювань або підрахунків, які дозволяють дати їм чисельну характеристику. Подібні ознаки прийнято називати кількісними. Чітку межу між якісними й кількісними ознаками провести неможливо: деякі кількісні можна описати якісно (наприклад, високий - карликовий, скоростиглий - пізньостиглий), якісні ж відмінності можна виразити кількісно (наприклад, різницю в забарвленні - кількістю пігменту). Проте якісний опис можливий лише у рідких випадках, коли ці відмінності досить різкі й між ними немає проміжних форм. Більшість господарсько-корисних ознак культивованих рослин і домашніх тварин відносять до кількісних, тому розуміння того, як спадкуються кількісні ознаки, є дуже важливим для селекційної роботи.
Кількісні ознаки, як правило, мінливіші, ніж якісні. Це залежить від двох причин. По-перше, спадкові відмінності особин за тією чи іншою кількісною ознакою зазвичай зумовлені взаємодією декількох пар полімерних генів, причому кожен ген досить суттєво впливає на розвиток даної ознаки. Певні комбінації полімерних генів, які визначають кількісну ознаку, зсувають ступінь прояву ознаки в позитивний бік, інші - у негативний. На відміну від цього, у випадку якісних ознак різниця між особинами визначається здебільшого лише однією, двома або, зрідка, трьома парами алелів. По-друге, кількісні ознаки зазвичай значно сильніше, порівняно з якісними, залежать від зовнішніх факторів. При цьому різні фактори часто можуть діяти на фено-типовий прояв генів різнонаправлено.
При вивченні спадкування кількісних ознак використовують певні статистичні параметри, за допомогою яких характеризують ступінь прояву таких ознак. Головні з цих параметрів - середнє арифметичне (x ), середнє квадратичне відхилення (о) і коефіцієнт мінливості (CV).
Ознаку, яку вивчають, вимірюють в усіх особин досліджуваної групи, отримані дані розбивають на довільну, але відносно невелику кількість класів, у кожному з яких об'єднані особини більш-менш схожі за значенням ознаки. Тобто створюють варіаційний ряд, який зручно зображувати у вигляді гістограми - стовпчиків з основами, які відповідають інтервалу, прийнятому для даного класу, і висотою, що відповідає кількості варіантів (тобто кількості особин, які належать до даного класу ). Аналогічно можна побудувати криву розподілу (рис. З.7).
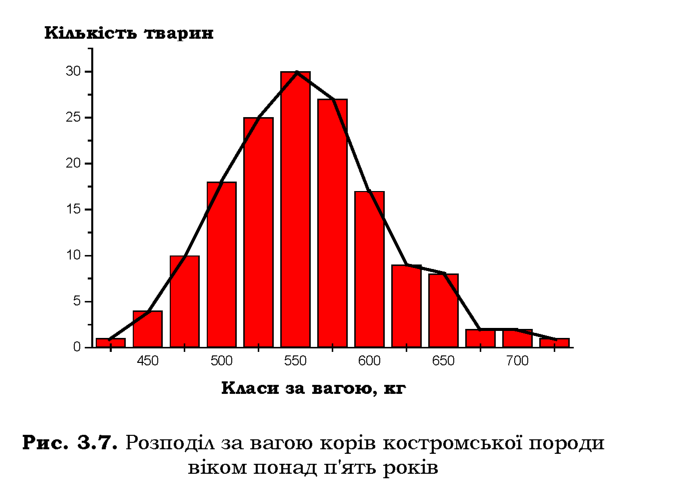
Після побудови варіаційного ряду знаходять середнє арифметичне (x ), яке дає уявлення про типове значення ступеня прояву ознаки в досліджуваній групі особин. Середнє арифметичне розраховують за формулою
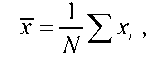
де х - значення для окремих особин; N - кількість особин.
Середнє арифметичне не дає уяви про ступінь мінливості ознаки в межах досліджуваної групи: варіаційні ряди, які мають однакове середнє арифметичне, можуть значно відрізнятися за мінливістю. Найуживанішою мірою мінливості (ширини розподілу ознаки) є середнє квадратичне відхилення (о), його розраховують за формулою
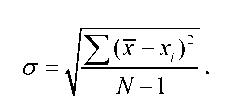
Середнє квадратичне відхилення дозволяє порівнювати мінливість однієї й тієї ж ознаки в різних групах особин. Проте цей показник не придатний для порівняння мінливості ознак, які характеризуються різними одиницями вимірювання, наприклад, мінливості ваги та зросту. У такому випадку розраховують коефіцієнт варіації (CV), що являє собою виражене у відсотках відношення середнього квадратичного відхилення до середнього арифметичного:
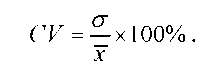
Потрібно зауважити, що не слід користуватися коефіцієнтом варіації там, де можна обійтися середнім квадратичним відхиленням, -у математичному відношенні останнє значно точніше характеризує мінливість ознаки.
Важливо розуміти також, що будь-який емпіричний варіаційний ряд є випадковою вибіркою, яка містить обмежену кількість варіантів, і тому не зовсім точно відображає характерну для даної популяції величину досліджуваної ознаки та 'її мінливість. Достовірність біометричних показників, що визначаються для емпіричної вибірки, сильно залежать від кількості особин, що входять до 'її складу: чим більша вибірка, тим ближчі ці показники до свого істинного значення.
Як правило, кількісні ознаки залежать від полімерних генів, що й зумовлює деякі особливості їхнього спадкування. При цьому в основі спадкової передачі кількісних ознак лежать такі самі явища розщеплення та перекомбінації генів, що й при передачі якісних. При схрещуванні особин, які відрізняються за кількісною ознакою, деякі нащадки у F2 можуть мати більше чи менше значення прояву ознаки, ніж вихідні батьки. Якщо обидва батьки несуть як гени, що збільшують це значення, так і ті, що зменшують його, то у F2 мають з'явитися нащадки, в яких концентрація плюс- або мінус-генів буде вищою, ніж у батьків. Припустимо, наприклад, що полімерні гени A, B і C збільшують прояв ознаки, а їхні алелі a, b і c зменшують його, і що генотип однієї батьківської форми ААВВсс, а іншої - aabbCC. Тоді серед особин F2 деякі матимуть генотип ААВВСС або aabbcc; у перших значення ознаки буде більше, а в других - менше, ніж у батьків. Така можливість розсунути межі спадкової мінливості кількісної ознаки має велике практичне значення. У багатьох випадках цим шляхом вдається виділити з F2 форми, які генотипово і фенотипово є ціннішими, ніж вихідні батьківські форми.
У найпростішому вигляді теорія полімерних генів припускає, що будь-яка кількісна ознака визначається взаємодією кількох адитивно діючих генів, які мають приблизно однаковий вплив на цю ознаку. Насправді полімерні гени часто різняться за "силою", з якою кожен із них визначає прояв даної ознаки: залежно від ступеня впливу на кількісну ознаку гени поділяють на сильні та слабкі, а також гени з проміжним впливом на прояв ознаки. Деякий ген із плейотропною дією може виступати як сильний щодо одних ознак і як слабкий -стосовно інших. Крім того, взаємовідносини полімерних генів можуть бути неадитивними й нерідко ускладнюються явищами комплемен-тарності або епістазу, а також наявністю слабкіших генів-модифікаторів, вплив яких складно враховувати.
Більш або менш точне визначення конкретного набору генів, які визначають прояв кількісної ознаки, є досить складним завданням. У практиці рослинництва і тваринництва постійно виникає потреба знати, якою мірою мінливість кількісної ознаки зумовлена генетичними причинами, а якою - модифікаціями, викликаними факторами зовнішнього середовища. Від розв'язання цієї проблеми залежить прогноз ефективності добору, що проводиться за даною ознакою. У таких випадках суттєву користь може принести обчислення коефіцієнта успадковуваності ознаки. Це може бути здійснено й без установлення конкретних генів, які визначають цю ознаку. Коефіцієнт успадковуваності є величиною, що показує, яка частка припадає на генетичну компоненту в загальній фенотиповій мінливості досліджуваної кількісної ознаки. Фенотипова мінливість ознаки характеризується середнім квадратичним відхиленням або квадратом цього відхилення - дисперсією (о2). Загальна фенотипова дисперсія (о2р) складається з дисперсії, що залежить від генетичної різноманітності особин (o2g), і дисперсії, зумовленої зовнішніми впливами (о2е). Коефіцієнт успадковуваності h2 визначається рівнянням:
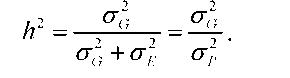
Існує декілька різних способів обчислення коефіцієнта успадкову-ваності. Найпростіший із них базується на зіставленні мінливості кількісної ознаки в генетично однорідній групі особин із мінливістю цієї ж ознаки в групі особин, узятої з гетерогенної популяції того самого виду. Генетично однорідну групу отримують у результаті того, що протягом багатьох поколінь розмноження здійснюється шляхом схрещування близькоспоріднених особин або самозапилення.
ХРОМОСОМИ ЯК ГРУПИ ЗЧЕПЛЕННЯ ГЕНІВ
Той факт, що гени містяться у хромосомах, які, власне, і передаються від батьків до нащадків, суттєво обмежує третій закон Менделя: незалежно одна від одної спадкуються хромосоми - великі групи генів (групи зчеплення). Зрозуміло, що це обмеження має зумовлювати відхилення від менделівських розщеплень у разі полігибридних схрещувань. Наприклад, при схрещуванні дигетерозигот AaBb, за умови, що гени А/а і B/b знаходяться в одній хромосомі, замість розщеплення 9 : 3 : 3 : 1 має бути отримано два фенотипові класи у відношенні 3 : 1 (рис. 3.8). Іншим засобом перевірити наявність генів в одній хромосомі, є аналізуюче схрещування AaBb х aabb: якщо два гени спадкуються незалежно, то утворюються чотири фенотипові класи нащадків у співвідношенні 1 : 1 : 1 : 1 (скористуйтесь схемою моногібридного аналізуючого схрещення, наведеною вище, і правилом добутку ймовірностей); якщо гени зчеплені в одній хромосомі, буде два фенотипових класи (AaBb, aabb) у рівному співвідношенні.
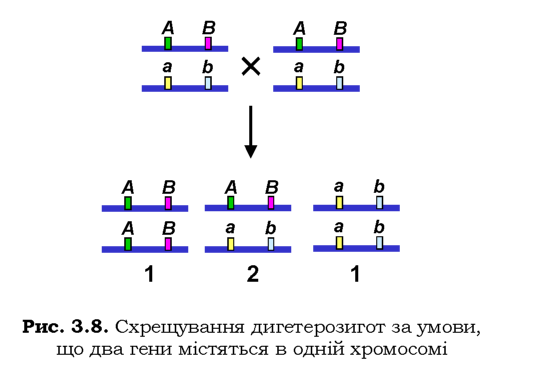
Спостереження такого типу та встановлення того факту, що кількість груп зчеплення дорівнює гаплоїдній кількості хромосом, дозволили свого часу Моргану (Thomas Hunt Morgan) і співробітникам отримати генетичні докази того, що гени розташовані у хромосомах (див. історичну довідку).
Проте, і це теж було встановлено групою Моргана, очікувані співвідношення на кшталт щойно наведених майже ніколи не реалізуються точно: обмін ділянками між гомологічними хромосомами при мейозі -кросинговер (див. розділ 1) - приводить до "перемішування" генів у межах гомологічних груп зчеплення (гомологічних хромосом) і, таким чином, частково відновлює незалежність передачі генів до нащадків.
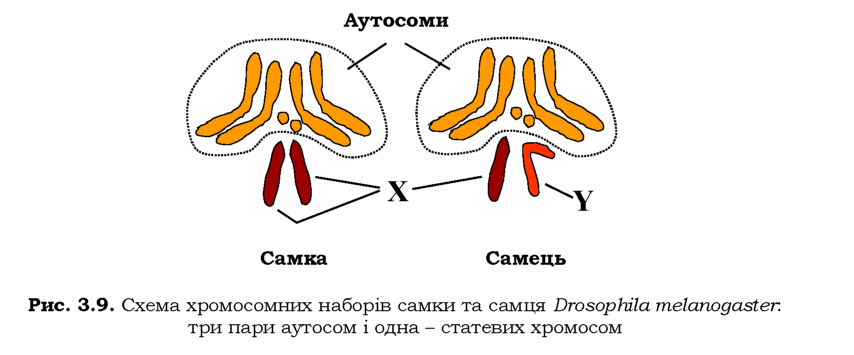
Останнє твердження потребує уточнення: кросинговер унаслідок гомологічної рекомбінації відбувається тільки в парах гомологічних хромосом, які є однаковими за морфологією та набором генів у обох статей, -так званих аутосом. У більшості організмів, які розмножуються статевим шляхом, одна пара хромосом - статеві - представлена двома него-мологічними типами (детальніше про генетику статі йдеться в розділі 6). Одна зі статей при цьому (гомогаметна) характеризується двома однаковими статевими хромосомами, інша (гетерогаметна) - двома різними (наприклад, у ссавців і деяких комах дві X-хромосоми визначають самку, X- і Y-хромосома - самця, рис. 3.9). Рекомбінація в парі негомологічних статевих хромосом є неможливою (або суттєво обмеженою), отже статеві хромосоми передаються як одне ціле від гетерогаметної статі до нащадка.
Спадкування ознак, зчеплених зі статтю
Суттєвою обставиною, яка також впливає на розподіл фенотипо-вих класів, є те, що негомологічні статеві хромосоми містять різні набори генів. Не слід думати, що це гени, які визначають статеві ознаки: більшість генів X-хромосоми людини та комах кодує ознаки, спільні для обох статей, тоді як Y-хромосома взагалі є практично інертною, оскільки в ній майже немає активних генів. Спадкування ознак, гени
яких розташовані в статевих хромосомах, називається спадкуванням., зчепленим зі статтю, і для нього спостерігаються суттєві відхилення від менделівських розщеплень.
Уперше таке спадкування досліджено Морганом на прикладі плодової мушки Drosophila melanogaster - об'єкта, що відіграв надзвичайно велику роль у розвитку генетики. Актуальним він залишається й досі.
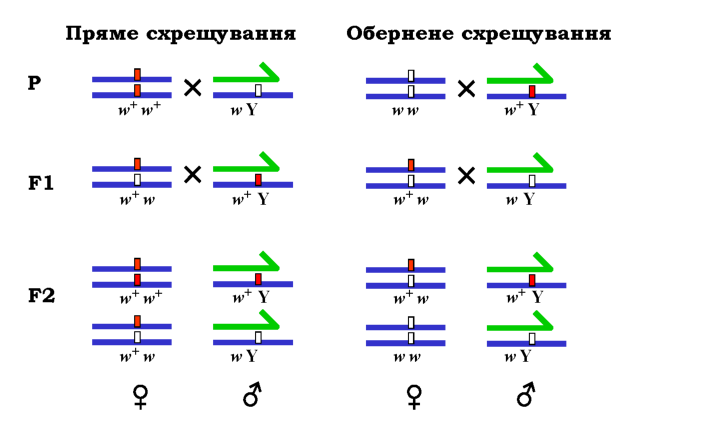
У Х-хромосомі дрозофіли знаходиться один із генів, від яких залежить розвиток червоного забарвлення ока ("нормальні" мухи - мухи дикого типу - мають червоні очі). Мутація цього гена перетворює його на мутантний рецесивний алель white (скорочене позначення w, відповідний алель дикого типу позначають як w+) - гомозигота за рецесивним алелем характеризується білими очима. Морган поставив два так звані реципрокні схрещування (рис. 3.10): самка з червоними очима х самець із білими (пряме схрещування), і навпаки - білоока самка х червоноокий самець (обернене схрещування).
 |
|
Рис. 3.10. Схема реципрокних схрещувань дрозофіл, які несуть рецесивну мутацію w (білі очі) і мають відповідний алель дикого типу w+ (червоні очі). Х-хромосому позначено синім кольором, Y-хромосому - зеленим |
У прямому схрещуванні нащадки першого покоління всі були червоноокі, тобто виконувався перший закон Менделя. Але в оберненому схрещуванні вже в першому поколінні спостерігалося розщеплення на червонооких і білооких таким чином, що всі самки були червоноокі, а самці - білоокі. Така картина спадкування, коли у ознаки батьків передаються нащадкам протилежній статі, називається крис-крос (criss-cross).
При схрещуванні особин першого покоління у F2 у прямому схрещуванні було отримано загальне розщеплення З : 1, але самки -всі червоноокі, самці - червоноокі та білоокі у співвідношенні 1 : 1. В оберненому схрещуванні відношення 1 : 1 спостерігалося як для самців, так і для самок.
Оскільки наведені розщеплення повністю корелюють із поведінкою статевих хромосом, описаний експеримент став свого часу одним із ключових доказів ролі хромосом у спадковості.
1-і • • ^ • • •
Генетичні наслідки обмінів ділянками між гомологічними хромосомами
Як уже йшлося, при схрещуванні дигетерозиготи AaBb із гомозиготою aabb мають утворитися два фенотипові класи з генотипами AaBb і aabb, якщо гени знаходяться в одній хромосомі, і чотири класи (АaBb, aabb, Aabb, aaBb) - якщо в різних. Проте, розглядаючи випадок зчеплення двох генів, слід узяти до уваги гомологічну рекомбінацію (розділ 1), яка відбувається під час мейозу при гаметогенезі та приводить до обміну гомологічними ділянками між гомологічними хромосомами - кросинговеру. Зрозуміло, що внаслідок кросинговеру в дигетерозиготного батька, крім гамет, які містять комбінації генів AB і ab, із певною імовірністю утворяться також гамети з комбінаціями Ab і aB. У результаті серед нащадків, крім двох генотипів AaBb і aabb, будуть знайдені також генотипи Aabb, aaBb.
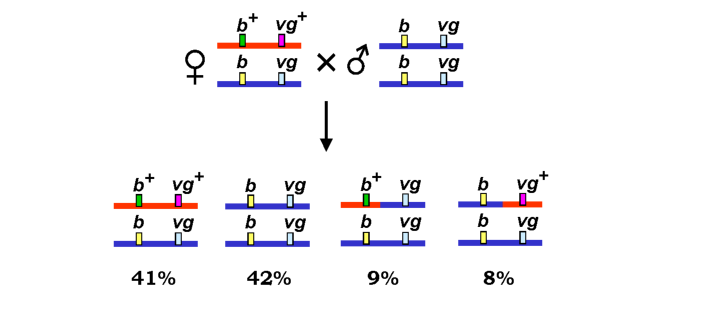
Саме таку ситуацію ілюструє рис. З.11, на якому наведено результати конкретного схрещування дрозофіл, здійсненого групою Моргана. У другій хромосомі дрозофіли присутні два гени, рецесивні алелі яких зумовлюють чорне забарвлення тіла (black або b, алель дикого типу - сіре тіло - позначається як b+) і редуковані крила (vestigial або vg, алель дикого типу - vg+). Аналізуюче схрещування дигетерозиготної самки з гомозиготним самцем приводить до розщеплення нащадків на чотири фенотипові класи. Більшість особин розподілено в рівному співвідношенні між двома класами: диким типом та мухами з чорним тілом і редукованими крилами - саме тими класами, що очікуються з огляду на зчеплення двох генів. Але 17 % особин представлено (також у рівному співвідношенні) двома фенотиповими класами, які мали б утворитися при незалежному спадкуванні. Ці два класи називають кросоверними - як показано на схемі рис. 3.11, вони утворилися внаслідок обміну ділянками між гомологічними хромосомами самки. Оскільки самець є подвійною гомозиготою, обмін ділянками його хромосом у випадку, що розглядається, не мав би генетичних наслідків, але слід зауважити, що в самців дрозофіл кросинговер при сперматогенезі взагалі не відбувається.
 |
|
Рис. 3.11. Схема схрещування самців дрозофіл, гомозиготних за двома рецесивними алелями, які містяться в одній хромосомі, із самками, гетерозиготними за цими генами |
Зрозуміло, що для двох хромосомних локусів перенесення внаслідок кросинговеру першого чи другого з них на гомологічну хромосому є рівноймовірними подіями. Відповідно, частоти двох кросоверних класів є практично однаковими (рис. 3.11). Частота кросинговеру (у нашому прикладі вона становить 9 + 8 = 17 %) визначається як відношення кількості гамет із обмінами між двома локусами до загальної кількості гамет. Звичайно, відносна кількість гамет різних типів визначається за результатами відповідних схрещувань.
Зрозуміло також, що чим більшою є відстань між двома хромосомними локусами, тим більше рекомбінаційних подій у більшій кількості точок може відбутися між ними. Це означає, що частота кросинговеру є мірою фізичної відстані між локусами, і у відсотках кросинговеру можна вимірювати відносну відстань між генами на хромосомі. Саме таким чином протягом довгого часу, а іноді й досі, здійснювали картування хромосом - установлювали лінійний порядок розміщення генів. Одиницею виміру відстані на таких картах є сантиморган (centiMorgan, cM): 1 сМ відповідає відстані, що забезпечує частоту кросинговеру в 1 %. Отже, відстань між генами black і vestigial становить 17 сМ.
Відносна відстань між генами, яку можна визначити за частотою кросинговеру, має верхню границю - при відстані 50 сМ і більше взагалі неможливо встановити, чи містяться гени в одній хромосомі. Припустимо, що відстань між двома генами дорівнює цій граничній величині - кросинговер відбувається із частотою 50 %. Тоді половина гамет, котрі продукує дигетерозигота (як на рис. З.11), є кросоверни-ми, - утворюється всього чотири класи гамет по 25 % у кожному.
І, відповідно, в аналізуючому схрещуванні буде отримано співвідношення фенотипових класів 1 : 1 : 1 : 1 - таке саме, що й при незалежному спадкуванні генів, розташованих у різних хромосомах.
Власне, у перемішуванні генів між двома гомологічними хромосомами, яке забезпечує (повністю або частково - залежно від фізичної відстані між генами) справедливість третього закону Менделя, і полягає основна біологічна роль кросинговеру. Справді, якщо б не було кросинговеру, то при схрещуванні полігетерозигот гомозигота за багатьма рецесивними алелями одночасно (n алелів, розташованих в одній хромосомі) утворювалася б в одному випадку із чотирьох (за другим законом Менделя, див. рис. З.8). Кросинговер забезпечує для багатьох пар генів практично незалежне спадкування, і ймовірність утворення такої гомозиготи зменшується на кілька порядків -до 1/4n (див. табл. З.1). З огляду на те, що більшість рецесивних алелів є мутантними, кросинговер, таким чином, різко підвищує частку життєздатних гетерозигот у наступному поколінні. Крім того, викликане кросинговером незалежне спадкування генів зумовлює створення великої кількості їхніх комбінацій у межах популяції, що сприяє, з одного боку, загальній генетичній стабільності популяції, з іншого -забезпечує певний еволюційний потенціал.
Кросинговер відбувається в усіх досліджених видів тварин і рослин, хоча наявність процесу та його частота може залежати від статі: наприклад, у самців дрозофіли, як уже згадувалося, і самок шовкопряда (обидві статі є гетерогаметними) кросинговер не відбувається; у людини рекомбінація здійснюється у два рази частіше при сперматогенезі у чоловіків, ніж при оогенезі у жінок. Частота кросинговеру залежить також від факторів зовнішнього середовища (наприклад, від температури для пойкілотермних організмів) і стадій індивідуального розвитку. Уздовж хромосоми частоти гомологічної рекомбінації розподілені нерівномірно: залежність частоти від фізичної відстані є лінійною в середніх частинах плечей хромосом, ближче до гетерохроматинових центромерних і теломерних зон кросинговер пригнічується. Існують також певні "гарячі точки", які характеризуються підвищеною частотою рекомбінації. Нарешті, як і будь-який інший біологічний процес, гомологічна рекомбінація контролюється певним набором генів, мутації яких також впливають на частоту кросинговеру.
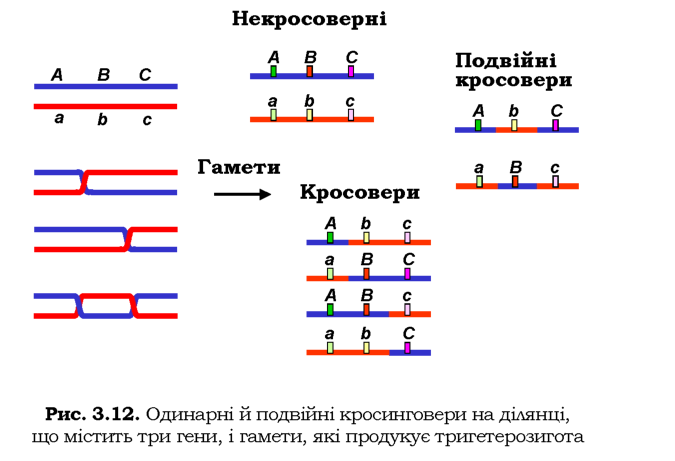
Кросинговер, що спостерігається між двома хромосомними локу-сами (як на рис. 3.11), є наслідком багатьох рекомбінаційних подій на ділянці, котра розділяє ці два гени: у генетичному аналізі детек-тується сумарний результат усіх таких подій, його можна формально розглядати як одинарний обмін ділянками. Термін "подвійний кросинговер" (потрійний або вищого порядку) відображає тільки той факт, що аналізуються генетичні наслідки рекомбінаційних подій на ділянці хромосоми, яка має три маркерні локуси (чотири для потрійного кросинговеру і т. д.).
Наприклад, якщо аналізувати рекомбінаційні події між двома ло-кусами А і С на рис. 3.12 (припустимо, що ділянка В не містить маркерного гена, який мав би фенотиповий прояв), то кросинговери на ділянках між А і В і між А і С будуть сприйматися як одинарний кросинговер десь на ділянці між А і С, а подвійне перехрестя, яке повертає локус С до вихідної хромосоми, узагалі не буде зафіксоване як рекомбінаційна подія. Якщо ж на ділянці В є ген, який можна аналізувати, то всі зазначені рекомбінаційні події будуть сприйматися окремо: вісім типів гамет, які продукує тригетерозигота на рис. 3.12, можна буде зафіксувати в аналізуючому схрещуванні (з гомозиготою за трьома рецесивними алелями abc/abc).
Результати такого гіпотетичного аналізуючого схрещування наведено в табл. 3.11. За частотою кросоверних особин відстань між ло-кусами А/а і В/b дорівнює 15 + 3 = 18 сМ (подвійний кросинговер також спричинює реципрокне переміщення В/ b до гомологічних хромосом, і, якщо б аналізували лише два локуси - А/а і В/b, - то ці подвійні перехрестя давали б внесок у частоту формально одинарного кросинговеру). Аналогічно, відстань між локусами В/b і С/с становить 26 + 3 = 29 сМ. Отже, відстань між А/ а і С/ с (вона дорівнює сумі
відстаней між А/а і В/b та між В/b і С/с) можна розрахувати як суму частот одинарних кросоверів, додавши до неї подвоєну частоту подвійних кросоверів 15 + 26 + 2 х 3 = 47 сМ.
|
Таблиця 3.11. Розщеплення в аналізуючому схрещуванні тригетерозиготи з рис. 3.12 |
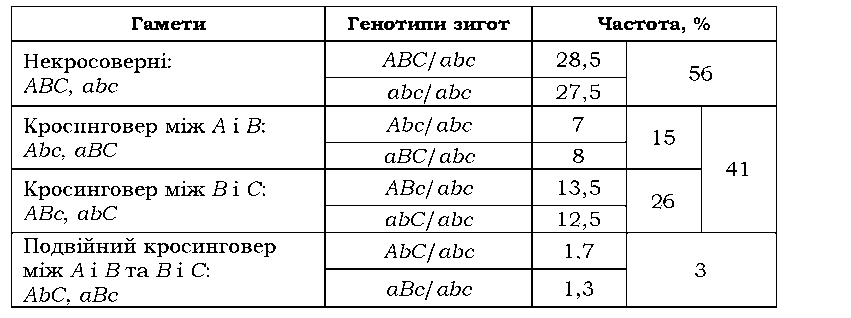 |
Слід зауважити, що в разі відсутності локусу В/b відстань між А/а і С/с була б оцінена як 18 + 29 - 2 х 3 = 41 сМ (табл. 3.11): подвійний кросинговер на ділянці між А/ а і С/ с знижує спостережувану частоту кросоверних форм. Узагалі, чим більша відстань між двома генами, тим
вища ймовірність подвійних (і вищого порядку) кросинговерів і тим менш точно можна визначити відстань. І навпаки, відстань визначається тим точніше, чим вона менша. Наприклад, в одному з класичних експериментів групи Моргана, визначалися відносні відстані між трьома генами, розташованими в Х-хромосомі дрозофіли: у (yellow - жовтий колір тіла ), w (ген white, що вже згадувався) та bi (bifid - вилкоподібні крила). Подвійні кросовери в цьому випадку взагалі не спостерігалися, частота кросоверів між генами у і bi (4,7 сМ) точно дорівнювала сумі частот кросоверів між генами у і w (1,2 сМ) та w і bi (3,5 сМ) - подібні спостереження і стали основою для формулювання концепції щодо лінійного розташування генів на хромосомі.
Повертаючись до нашого прикладу на рис. 3.12, треба зазначити, що наведена в табл. 3.11 частота подвійних кросоверів (3 %) суперечить припущенню про повну незалежність між рекомбінантними подіями на сусідніх ділянках хромосом. Якщо б такі події на ділянках між ло-кусами А/а і В/b та В/b і С/с були незалежними, то частота подвійних кросоверів (результату двох одночасних подій на цих ділянках) мала б дорівнювати добутку частот відповідних одинарних кросинговерів (за правилом добутку ймовірностей): 0,18 х 0,29 = 0,052 (5,2 %). Зазначимо, що в якості частоти одинарних подій слід брати саме сумарну частоту кросинговеру (наприклад, 0,15 + 0,03 = 0,18): важливим є те, з якою частототою алелі В/ b обмінялися місцями (див. табл. 3.11), і неважливо, чи повернулися при цьому алелі С/с на "своє місце" внаслідок подвійного кросинговеру.
Отже, обмін між локусами на одній ділянці впливає на такий обмін на сусідніх ділянках - явище, яке називають інтерференцією. У нашому прикладі (і це типова ситуація) інтерференція є позитивною - кросинговери на сусідніх ділянках заважають один одному. Зрозуміло, що інтерференція є тим суттєвішою, чим меншою є відстань між двома ділянками. Іноді спостерігається також негативна інтерференція, коли відбувається взаємна стимуляція рекомбінаційного процесу на двох або більше сусідніх ділянках. Проте насправді негативна інтерференція відображає не підвищення частоти подвійних кросинговерів, а є наслідком конверсії гена (див. нижче).
Для оцінки відповідності між очікуваною на підставі незалежності окремих рекомбінаційних подій частотою подвійного кросинговеру h0 та частотою h, що спостерігається, використовують коефіцієнт коінцеденції
С = h/ha. Тобто, у нашому прикладі С = 0,0З0/0,052 = 0,58, величина інтерференції становить І = 1 - С = 0,42.
З наведених оцінок зрозуміло, що інтерференція частково компенсує вплив подвійного кросинговеру на визначення відстані між двома генами. Справді, якщо б дві рекомбінаційні події були незалежними, і частота подвійного кросинговеру була б 5,2 %, то частота кросинговеру між локусами А/а і С/с (оцінена у відсутності В/b) становила б 18 + 29 - 2x5,2 = З6,6 сМ.
 |
|
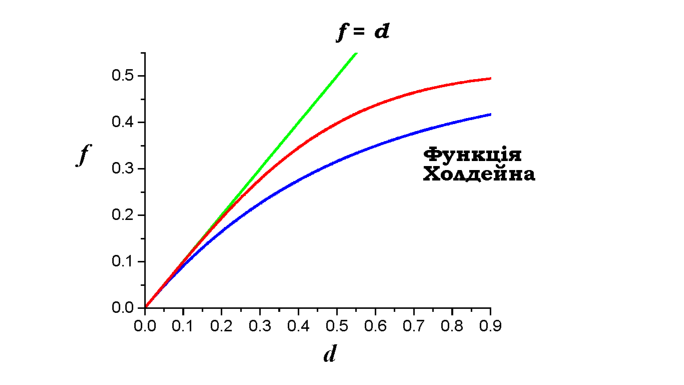
Рис. 3.13. Реальна залежність частоти кросинговеру між двома локусами f від відстані d (червона крива) порівняно з двома ідеалізованими залежностями |
Розрахована частота кросинговеру f між двома локусами як результат одинарних і множинних перехресть у припущенні про незалежність окремих рекомбінаційних подій описується функцією Холдейна (John Haldane):
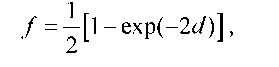
де d - фізична відстань між двома локусами в одиницях імовірності (змінюється від 0 до 1). На малих відстанях exp(-2d) ~ 1 - 2d і f ~ d (частота кросинговеру є прямо пропорційною фізичній відстані, практично це справедливо для відстаней до ~10 сМ), на великих відстанях f наближається до 1/2 - максимально можливої частоти кросинговеру. Унаслідок інтерференції реальна залежність частоти від фізичної відстані займає проміжне положення між функцією Холдейна та прямою f = d (рис. З.1З).
Явище конверсії гена має тісний зв'язок із процесом гомологічної рекомбінації - незалежно від того, чи відбувся обмін ділянками між гомологічними хромосомами (див. розділ 1). Як показано на рис. 3.14, унаслідок конверсії, що відбулася з перетворенням алеля B на b або навпаки, гетерозигота Bb може продукувати гамети двох типів у співвідношенні 3 : 1. У генетичному аналізі за допомогою схрещувань такі гамети можуть формально розглядатися як подвійні кросовери (порівн. рис. 3.12), що характеризуються негативною інтерференцією - висока частота "подвійних кросоверів" за відсутності кросоверних форм між локусами А/а і С/с.
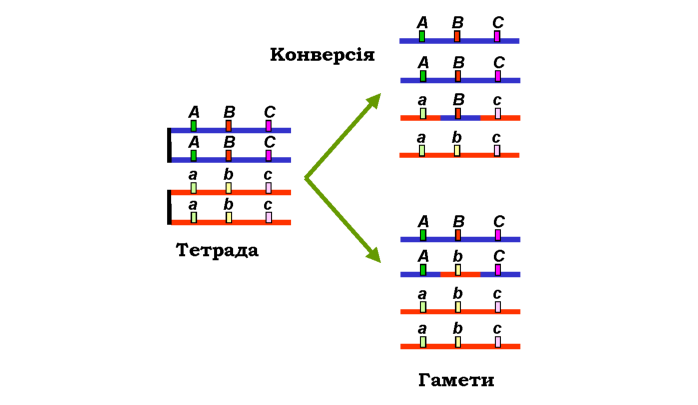 |
|
Рис. 3.14. Тетрада у профазі І мейозу та два варіанти набору гамет, які вона продукує у випадку конверсії гена B/ b між двома гомологічними хроматидами |
Слід відмітити, що конверсія гена створює для диплоїдних організмів можливість відновлення генетичної інформації, яка виявилася пошкодженою внаслідок мутації в одній із двох гомологічних хромосом (див. також розділ 5).
Гомологічна рекомбінація - досить точний процес, який відбувається між гомологічними ділянками ДНК (див. розділ 1). У результаті, зазвичай при кросинговері, як і в усіх попередніх прикладах, відбувається обмін однаковими за довжиною ділянками, що містять однакову кількість генів.
Але іноді спостерігається рекомбінація між гомологічними ділянками, які займають різне положення на хромосомі, унаслідок чого здійснюється обмін неоднаковими за довжиною ділянками - нерівний кросинговер. Як показано на рис. З.15, такий обмін зумовлює дуплікацію ділянки в одній хромосомі та делецію - в іншій. Таким чином, нерівний кросинговер є однією з причин хромосомних перебудов (див. розділ 4).
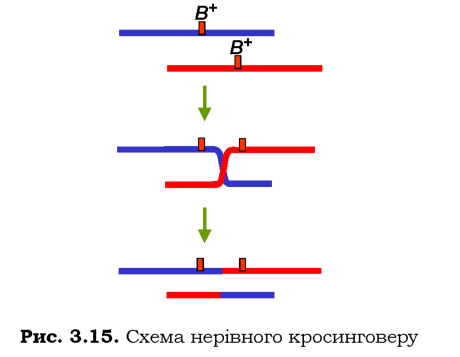
Саме така ситуація спостерігається для хромосомного локусу, розташованого в Х-ромосомі дрозофіли, який впливає на кількість фасеток ока: мухи дикого типу містять один такий локус (позначається як В+), дуплікація локусу (яка позначається як мутація Вar або В) приводить до зниження кількості фасеток. Повторення нерівного кросинговеру між одинарним і дуплікованим локусами веде до триплікації -мутація ВВ, очі мають ще меншу кількість фасеток.
Усі розглянуті приклади стосуються кросинговеру, що здійснюється під час мейозу при гаметогенезі. Але кросинговер може також відбуватися в соматичних клітинах - під час мітозу на стадії утворення чотирьох хроматид. Зокрема, такі випадки описано для дрозофіли. Так, самка, гетерозиготна за алелями локусу w (wm - очі кольору корала та w -білі очі) має очі рожевого кольору. Якщо личинки, що розвиваються, піддати рентгенівському опроміненню, то під час мітозу може відбутися обмін ділянками між несестринськими гомологічними хроматидами. У результаті в одній із дочірніх клітин опиняться дві хромосоми з але-лем w, а в іншій - дві з алелем wm. Перша клітина при наступному розмноженні дасть початок розвитку клону ідентичних клітин - білої плями на тлі рожевого ока, друга - зумовить появу темно-червоної плями.
Мітотичний кросинговер є наслідком точної репарації дволанцю-гових розривів під час реплікації за механізмом гомологічної рекомбінації (див. рис. 1.25). Після завершення процесу репарації та розділення структури Холідея, яка утворилася між двома дочірніми молекулами ДНК, ці молекули можуть обмінятися своїми ділянками. Для одноклітинних еукаріотів мітотичний кросинговер, напевно, є важливим засобом забезпечити рекомбінацію генетичного матеріалу при вегетативному розмноженні.
Контрольні запитання і завдання
1. Сформулюйте три закони Менделя та поясніть їх, виходячи з молекулярних і цитологічних механізмів спадковості.
2. Що таке генотип, фенотип, фенотиповий радикал, гомозигота, гетерозигота?
3. Яким чином зазвичай виникають рецесивні алелі?
4. Яка різниця між повним домінуванням, неповним домінуванням і кодомінуванням?
5. Розрахуйте розщеплення від схрещування дигетерозигот АаВв за умови неповного домінування для гена Аа.
6. Розрахуйте значення х2 для випадку випадіння монети 25 разів орлом і 32 рази - решкою. Чи виправдовується нульова гіпотеза?
7. Дайте визначення комплементарності, епістазу та полімерії при взаємодії неалельних генів.
8. Чим відрізняються підходи до вивчення кількісних і якісних ознак?
9. Опишіть статистичні параметри, за допомогою яких характеризують ступінь прояву кількісних ознак? Поясніть зміст коефіцієнту спадковості.
10. Скільки груп зчеплення генів існує у дрозофіли?
11. Запишіть схему та результати схрещування самки дрозофіли з жовтим тілом (зчеплена зі статтю рецесивна мутація y) із самцем дикого типу (чорне тіло, y+). Яке розщеплення буде реалізовано від схрещування нащадків (у другому поколінні)? Проаналізуйте також відповідне обернене схрещування.
12. Як визначається частота кросинговеру? В яких одиницях вимірюють відстань між генами на основі встановлення цієї частоти?
13. За якої умови є можливим дослідження подвійного кросинговеру?
14. Що таке інтерференція та який вплив вона має на частоту кросинговеру? Як визначається коефіцієнт коінцеденції?
15. Що таке конверсія гена й чому вона відбувається?
16. З яких причин відбувається нерівний кросинговер і які наслідки він має для структури гомологічних хромосом?
Автор: admin от 18-07-2013, 14:35, Переглядів: 67787