Мінливість генетичного матеріалу
Однією із загальних властивостей біологічних систем є їхня консервативність - здатність відновлювати генетичний матеріал і передавати його нащадкам у майже незмінному вигляді. Молекулярна машинерія реплікації та репарації ДНК (розділ 1) забезпечує надзвичайно високу точність відновлення генетичної інформації - рівень помилок становить ~10-10 нуклеотидних замін, тобто менше однієї заміни на еукаріотичний геном при одному подвоєнні ДНК. Зрозуміло, що абсолютна точність відтворення ДНК є неможливою, і мінливість генетичного матеріалу є також важливим аспектом його існування.
Мінливість можна визначити як здатність генетичного апарату до змін, які зумовлюють фенотипові відмінності між особинами одного виду в ряду поколінь або в межах одного покоління. Мінливість може бути спричинена змінами геному (спадкова мінливість) або виникати в результаті зміни експресії генів за дії факторів навколишнього середовища протягом індивідуального розвитку (неспадкова мінливість).
Спадкова мінливість може бути зумовлена утворенням або нових варіантів послідовностей нуклеотидів ДНК (мутаційна мінливість), або нових комбінацій уже існуючих послідовностей, які виникають за рахунок рекомбінації та випадкового розподілу хромосом у мейозі (комбіна-тивна мінливість). Механізми й наслідки комбінативної мінливості описано в розділах 1 і 3, а в цьому розділі увагу буде зосереджено саме на мутаційній мінливості. Слід розрізняти також власне мутаційну мінливість, яка приводить до випадкових змін генетичних програм (саме про таку мінливість ітиметься нижче), і запрограмовані зміни генетичного матеріалу в певних геномних зонах або іноді в масштабі цілого геному, приклади яких розглядатимуться в розділах 5 і 6.
Мутації - це незапрограмовані, випадкові та стабільні (такі, що залишаються "назавжди" та спадкуються) зміни в структурі ДНК, які з'являються або в результаті дії пошкоджуючих чинників, або як результат помилок систем реплікації, репарації чи рекомбінації. Мутація, яка виникла в соматичних клітинах (соматична мутація), спадкується тільки в ряду клітинних поколінь, а та, що в статевих (генеративна м,утація) - передається наступним поколінням нащадків. Крім мутацій, котрі виникають у ядерному геномі (ядерні мутаціі), зміни можуть відбуватися також у ДНК мітохондрій і хлоропластів - це цитоплазматичні мутації.
Мутаційні зміни можуть охоплювати декілька нуклеотидів молекули ДНК, великі за довжиною послідовності та цілі набори хромосом. Відповідно, розрізняють точкові, хромосомні та геномні мутації.
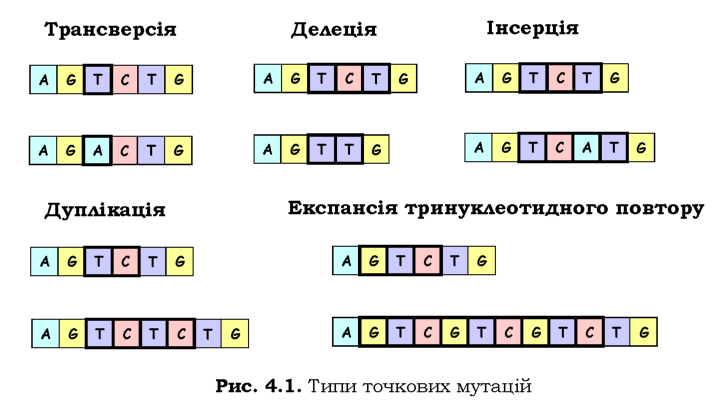
Зміну послідовності ДНК, обмежену лише одним чи декількома нуклеотидами, називають точковою мутацією. Така мутація може являти собою заміну одного нуклеотиду на інший. Заміна, в результаті якої замість пурину (А, G) включається піримідин (T, C) або навпаки називається трансверсією. Заміна пурину на пурин ^^G) і, відповідно, піримідину на піримідин (T^-C), називається транзицією. Транзиції та трансверсії, які відбулися в кодуючій частині гена, можуть зумовити заміну амінокислоти в складі білка - у такому випадку мутацію називають міссенс-мутацією (missense), або несинонімічною нуклеотидною заміною. Унаслідок виродженості генетичного коду (див. розділ 2) заміна нуклеотиду може не змінити змісту кодона. Така нуклеотидна заміна називається сеймсенс-мутацією (samesense), або синонімічною мутацією. При утворенні в результаті трансверсії чи транзиції стоп-кодона (беззмістовного щодо амінокислот) нуклеотидну заміну називають нонсенс-мутацією (nonsense).
Іншими двома типами точкових мутацій є вставка (інсерція) або випадіння (делеція) одного чи декількох нуклеотидів. Інсерція або делеція не кратної трьом кількості нуклеотидів у кодуючій частині гена приводить до зсуву рамки зчитування. Якщо вставка повторює послідовність, яка присутня поблизу місця інсерції, то її називають дуплікацією, а багаторазовий повтор декількох нуклеотидів - експансією повторів. Усі розглянуті типи точкових мутацій схематично зображено на рис. 4.1.
Хромосомні мутації (хромосомні перебудови або хромосомні аберації) - це порушення в нормальній морфології хромосом, спостережувані на стадії метафази або телофази мітозу, коли можна розрізняти окремі хромосоми. Отже, хромосомними мутаціями є такі великомасштабні зміни послідовностей ДНК (від 1 млн пар основ і більше), які можна ідентифікувати за допомогою оптичного мікроскопа. Здебільшого під хромосомними абераціями розуміють будь-які порушення морфології хромосом - у тому числі такі, що унеможливлюють наступний поділ клітини або взагалі є несумісними з життям і тому не спадкуюються. Хромосомні мутації - це частина хромосомних аберацій, які успадковуються дочірніми клітинами.
Хромосомні перебудови можуть бути внутрішньохромосомними (відбуваються в межах однієї хромосоми) і міжхромосомними (перебудови, що охоплюють дві різні хромосоми).
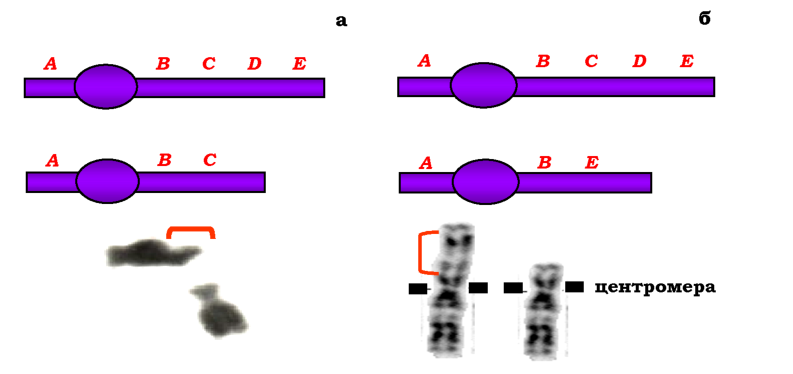
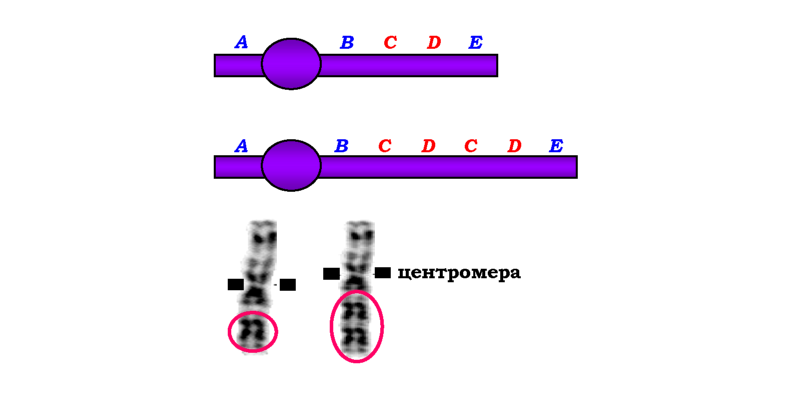
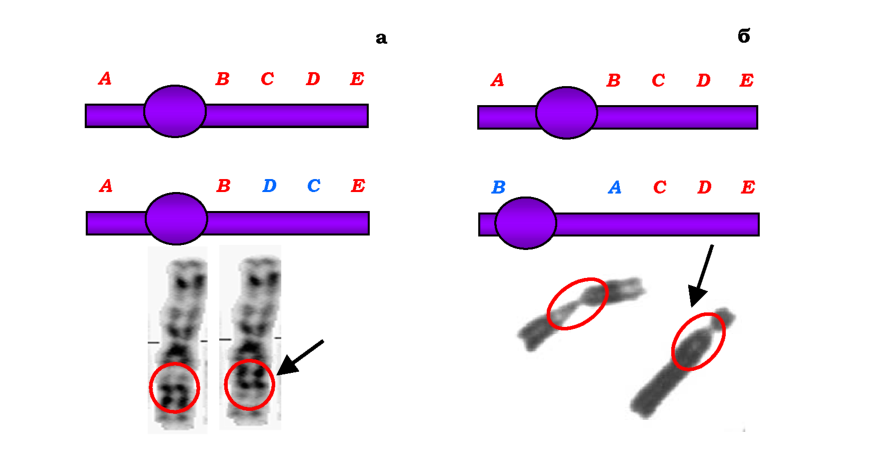
Одним із типів внутрішньохромосомних перебудов є делеції. Розрізняють термінальні делеції (дефішенсі - втрати кінцевих ділянок хромосом) та інтерстиціальні - втрати внутрішніх частин хромосом (рис. 4.2). До внутрішньохромосомних перебудов також належать дуплікації (двократні повтори певного сегменту хромосоми, рис. 4.3), ампліфікації (багаторазові повтори сегмента хромосоми) та інверсії -повороти ділянки хромосоми на 180°. Залежно від того, залучає інверсія область центромери чи ні, розрізняють відповідно перицентричні та парацентричні інверсії. Перші можуть значно змінювати морфологію хромосоми, а другі не приводять до зміни морфологічного типу хромосоми й детектуються лише за допомогою методів диференцій-ного забарвлення - методів, які дозволяють візуалізувати певні більш або менш інтенсивно забарвлені ділянки, розподіл яких є специфічною ознакою кожної хромосоми (рис. 4.4).
 |
|
Рис. 4.2. Схеми й фото термінальної (а) та інтерстиціальної (б) делецій. Овали на схемах позначають центромери. На фото наведено хромосоми людини, делеговані ділянки вказано квадратними дужками |
 |
|
Рис. 4.3. Схема та фото дуплікації. На фото - хромосоми людини, дуплікований фрагмент обведено червоним |
 |
|
Рис. 4.4. Схема та фото парацентричної (а) і перицентричної (б) інверсій. На фото - хромосоми людини, ділянки інверсій обведено червоним, хромосоми з інверсіями вказано стрілкою |
До міжхромосомних перебудов відносять інсерції та транслокації. Інсерція - це вставка ділянки однієї хромосоми всередину іншої (маються на увазі негомологічі хромосоми), яка супроводжується де-лецією в першій хромосомі.
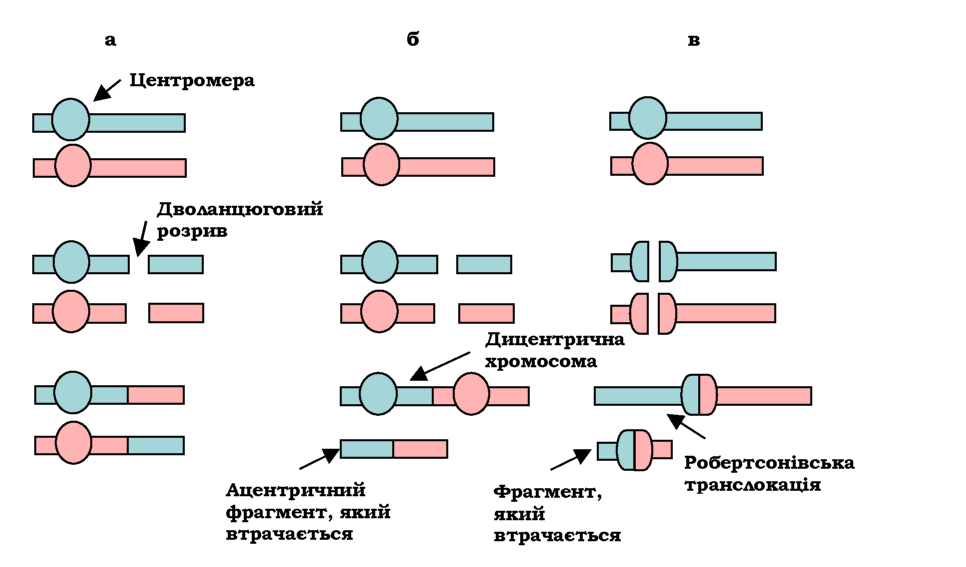
Обмін ділянками між негомологічними хромосомами називається транслокацією. Розрізняють реципрокні та нереципрокні транслокації. Взаємний обмін ділянками двох негомологічних хромосом - це реципрокні транслокації (рис. 4.5). При цьому можуть об'єднатися дві центромерні області різних хромосом, тоді обмін веде до утворення дицентричної хромосоми (рис. 4.5, б). Така хромосома є прикладом перебудови, що не спадкується, - це хромосомна аберація, яка не є мутацією. Різновидом реципрокних транслокацій є роберт-сонівські транслокації - об'єднання двох акроцентричних хромосом (після дволанцюгового розриву в центромерних зонах) у мета- чи субметацентричну (рис. 4.5, в). При цьому кількість хромосом зменшується на одну: тільки одна половина кожної центромери містить елементи послідовності, важливі для утворення кінетохору під час мітозу, відповідно, фрагмент, що утворився шляхом з'єднання інших половин, втрачається.
 |
|
Рис. 4.5. Реципрокні транслокації: без утворення дицентричної хромосоми (а); з утворенням дицентричної хромосоми (б); робертсонівська транслокація (в) |
При нереципрокних транслокаціях, на відміну від інсерцій, ділянка однієї хромосоми приєднується до кінця іншої (за умови порушення теломерної зони цієї іншої хромосоми).
Три типи геномних мутацій - гаплоїдія, поліплоїдія та анеуплоїдія -широко розповсюджені у тваринному й рослинному світі. Гаплоїдія -це зменшення вдвічі диплоїдного набору хромосом. Зворотним явищем є поліплоїдія - кратне гаплоїдному збільшення кількості хромосом. Клітина з трьома гаплоїдними наборами хромосом називається триплоїдною, чотирма - тетраплоїдною і т. д. Поліплоїдія може бути зумовлена або кратним збільшенням власних для даного виду хромосом (автополіплоїдія), або виникати за рахунок гібридизації, тобто об'єднання геномів різних видів (аллополіплоі'дія).
Анеуплоїдія - це не кратна гаплоїдному набору зміна кількості хромосом. Найчастіше вона виявляється у збільшенні або зменшенні кількості копій однієї хромосоми, рідше - декількох. Анеуплоїдна клі-
тина (чи організм), яка містить одну додаткову хромосому, має назву трисомік. Утрата однієї хромосоми приводить до моносомії, двох гомологічних хромосом - до нулісомії.
МОЛЕКУЛЯРНІ МЕХАНІЗМИ МУТАЦІЙНОЇ МІНЛИВОСТІ
Джерелом мутаційних змін є перебудови послідовності нуклеоти-дів ДНК, які виникають у результаті хімічних модифікацій молекули, таутомеризації азотистих основ, переміщення мобільних елементів (див. розділ 6), інтеграції чужорідної, (наприклад, вірусної) ДНК і помилок під час реплікації та репарації. Пошкодження ДНК не є власне мутаціями, а лише передмутаційними змінами, які можуть бути або виправлені системами репарації, або зафіксовані в ДНК у вигляді мутацій. Тобто мутація є такою зміною послідовності ДНК, яка залишилась після репарації та наступної чергової реплікації.
Пошкодження ДНК можуть виникати внаслідок як впливу продуктів нормальної життєдіяльності клітини, так і дії зовнішніх факторів середовища. Ендогенні та екзогенні фактори, здатні пошкоджувати ДНК, називають мутагенами, а процес утворення мутацій - мутагенезом. Мутагенез, який відбувається у природних умовах, конкретні причини якого, як правило, важко ідентифікувати, - це спонтанний мутагенез. Якщо мутації викликаються штучно (при використанні мутагенних факторів у експериментах), кажуть про індукований мутагенез. Молекулярні механізми виникнення пошкоджень ДНК і фіксації їх у вигляді мутацій принципово не відрізняються для обох типів мутагенезу.
Пошкодження ДНК, що виникають у процесі життєдіяльності клітини
Поява передмутаційних змін генетичного матеріалу в клітині унаслідок метаболічних процесів є нормальним явищем. Кількість спонтанних пошкоджень ДНК, що виникають у одній клітині людини за добу, оцінюється в 104-106. Переважна частина цих пошкоджень у нормі видаляється системами репарації, і тільки невелика кількість залишається у вигляді мутації.
Найпоширенішими передмутаційними пошкодженнями ДНК є втрати азотистих основ (утворення апуринових або апіримідинових сайтів), хімічні модифікації основ, ковалентні зшивання ДНК-ДНК і ДНК-білок, одно- та дволанцюгові розриви цукрофосфатного остова ДНК. Ці пошкодження виникають здебільшого в реакціях гідролізу (хімічні реакції з водою), реакціях з активними радикалами окси-гену та пероксидними радикалами, а також унаслідок метилування (алкілування) основ.
Гідроліз глікозидного зв'язку між азотистою основою та дезоксирибозою (див. рис. 1.1) приводить до видалення азотистої основи й появи в цьому місці апуринового чи апіримідинового сайта (АП-сайта). За добу в клітинній ДНК утворюється близько 10 тис. таких АП-сайтів. Неправильна репарація цих пошкоджень може зумовити нуклеотидні заміни (транзиції чи трансверсії). Відсутність репарації стане причиною того, що під час наступного реплікаційного циклу напроти АП-сайта у складі матричного ланцюга в ланцюзі, що синтезується, буде вставлено довільний нуклеотид - з імовірністю З/4 він виявиться не тим, що мав би стояти в цьому місці, тобто виникне точкова мутація типу нуклеотидної заміни. Крім того, ДНК-полімеразний комплекс може "проскочити" АП-сайт у складі матриці, наслідком чого буде делеція нуклеотиду в складі ланцюга, що синтезується.
Нерепаровані АП-сайти можуть також перетворюватися на одно-ланцюгові розриви. Накопичення одноланцюгових розривів, у свою чергу, приводить до розривів дволанцюгових (коли два одноланцюго-ві розриви розташовані на невеликій відстані та на різних ланцюгах), що може бути причиною утворення різних типів хромосомних аберацій. Одноланцюгові розриви ДНК можуть виникати також у результаті прямого гідролізу фосфодіефірного зв'язку.
При гідролізі екзоциклічних аміногруп азотистих основ виникає дезамінування основ (до 500 пошкоджень на клітину за добу). Так, у результаті дезамінування цитозин перетворюється на урацил (див. рис. 1.2), а 5-метилцитозин - на тимін. Обидві основи, що з'явилися внаслідок таких перетворень, комплементарні аденіну, а отже, у разі відсутності репарації дезамінування може зумовити нуклеотид-ну заміну (стабільну заміну пари GC на AT-пару при реплікації).
Оксидативні пошкодження ДНК виникають в результаті хімічних реакцій дезоксирибози та азотистих основ із вільними радикалами оксигену або пероксидними радикалами. Джерелом радикалів є процеси дихання клітини. Найсуттєвіші оксидативні пошкодження ДНК -це утворення 8-оксигуаніну (приєднання оксигену до восьмого атома кільця, див. рис. 1.2), комплементарного тиміну, і 2-оксиаденіну, комплементарного цитозину. Продуктами реакцій з вільними радикалами є також одноланцюгові розриви та зшивання ДНК-ДНК або ДНК-білок, які можуть бути причинами хромосомних аберацій.
Ще одним механізмом виникнення передмутаційних пошкоджень ДНК є метилування основ по атомах, які в нормі не піддаються цій модифікації. Помилки метилування викликають появу таких суттєвих пошкоджень ДНК: утворення 7-метилгуаніну, 3-метиладеніну та О6-ме-тиладеніну. 7-Метилгуанін і 3-метиладенін перешкоджають нормальному проходженню реплікації, унаслідок чого відбувається утворення однолацюгових прогалин (ділянок недореплікації) напроти модифікованих нуклеотидів. О6-метиладенін є комплементарним цитозину й може бути причиною транзицій.
Помилки реплікації та репарації
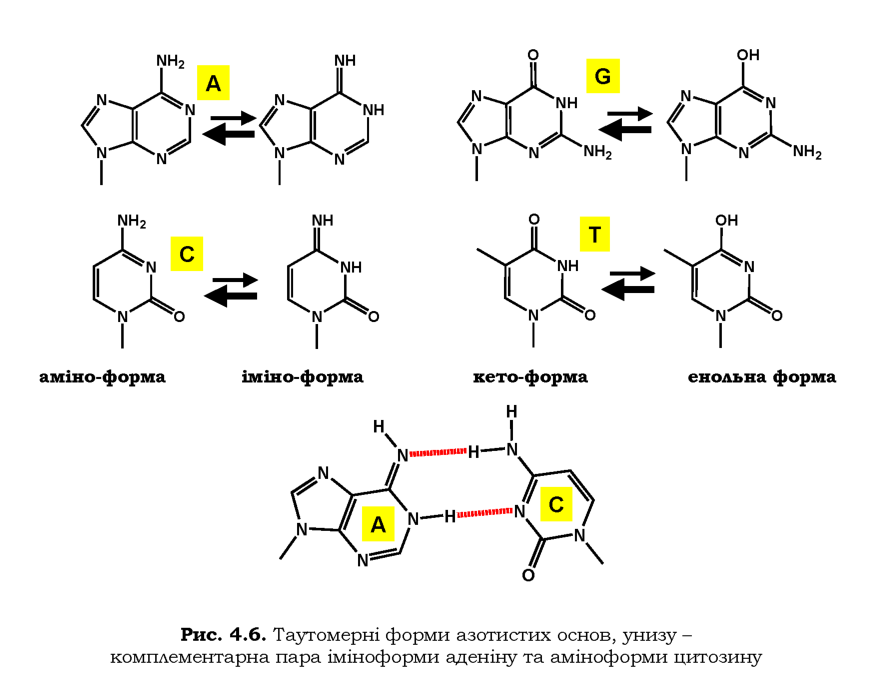
Помилкове включення нуклеотидів під час реплікації є досить вагомою причиною виникнення точкових мутацій і хромосомних перебудов. Утворення некомплементарних пар нуклеотидів (місметчів) під час реплікації відбувається з частотою 1 на 10 тис. нуклеотидних пар. Основною причиною помилкового приєднання нуклеотидів під час реплікації є таутом.ерш азотистих основ. Спонтанні перебудови електронних систем гетероциклів приводять до існування кожної основи у вигляді двох таутомерних форм: аміно- чи іміноформи для A, C; енольної чи кетоформи для G, T (рис. 4.6). Рівновага зсунута в бік аміно-та кетоформ, які й присутні у складі подвійних спіралей (див. також рис. 1.2) і для яких реалізуються правила комплементарності A-T, G-C. Але спарювання основ підпорядковується іншим правилам для мінорних таутомерних форм: наприклад, іміноформа А та аміноформа С утворюють між собою два водневі зв'язки (рис. 4.6), що може відбутися під час впізнання матриці черговим нуклеотидом при реплікації. Аналогічно, енольна форма тиміну є комплементарною гуаніну. У результаті швидкого повернення до мажорної таутомерної форми, у складі ДНК залишиться некомплементарна пара нуклеотидів. Якщо система редагування помилок під час синтезу ДНК і потім система репарації місметчів (див. розділ 1) не спрацює, у наступному репліка-тивному циклі така некомплементарна пара зафіксується у вигляді мутації в одній із двох дочірніх молекул.
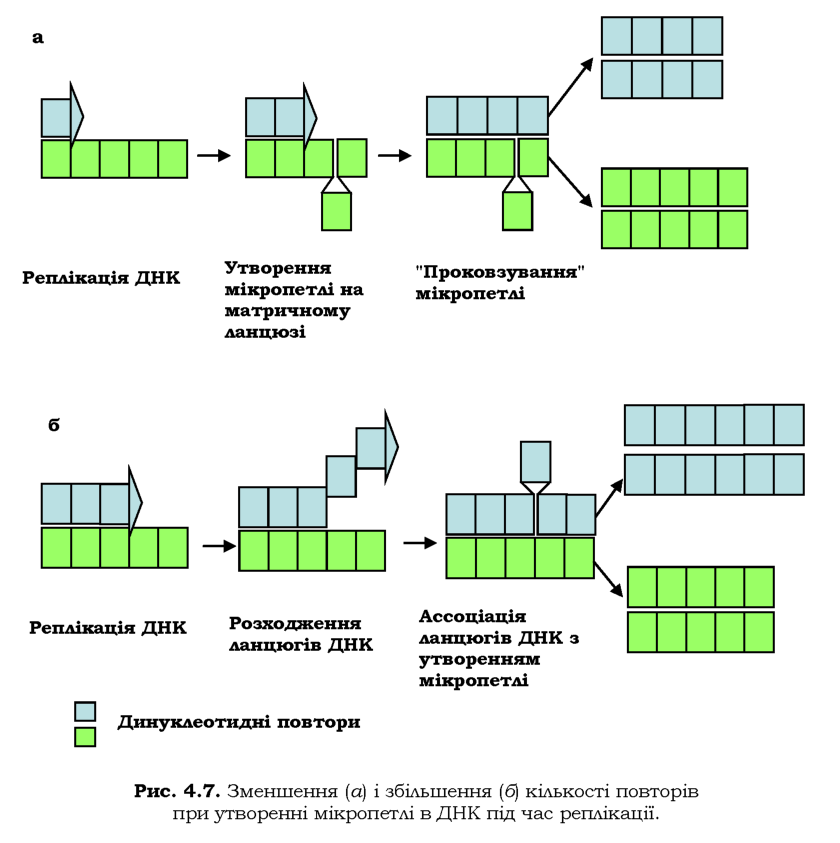
На ділянках мікросателітних тандемних повторів (повторів елементів послідовності довжиною 1-15 пар основ, розділ 6) спостерігається специфічна помилка ДНК-полімеразного комплексу - проковзування (slippage) ДНК-полімерази. На ділянці матричного або ново-синтезованого ланцюгів ДНК інколи відбувається утворення мікро-петель або мікрошпильок за рахунок внутрішньоланцюгових комплементарних взаємодій. У випадку появи мікропетлі на матричному ланцюзі ДНК дочірній ланцюг буде коротший на кілька нуклеотидів, і отже, після наступного раунду реплікації буде спостерігатися делеція (рис. 4.7, а). Якщо така мікропетля утворюється в дочірньому ланцюзі, кількість нуклеотидів у ньому збільшиться, що приведе до вставки одного або декількох повторів (рис. 4.7, б).
Мутації виникають не тільки внаслідок недостатньо ефективної репарації - деякі процеси репарації ДНК самі є причинами мутацій. Насамперед це стосується неточних систем репарації: SOS-репарації, яка зумовлює неточний синтез ДНК у разі великої кількості пошкоджень, що викликає ще більше зростання мутацій, і системи репарації дволанцюгових розривів за рахунок негомологічного з'єднання кінців NHEJ (див. розділ 1). Саме NHEJ вважається основною причиною реалізації хромосомних перебудов, оскільки забезпечує з'єднання кінців будь-яких молекул ДНК.
Механізми виникнення поліплоїдій і анеуплодій
Кількісні аномалії хромосом пов'язані з порушенням процесів, які забезпечують розходження хромосом (хроматид) у мітозі та мейозі. Причиною помилок розподілу хромосом можуть бути порушення контролю клітинного поділу, дефекти в центромерній області хромосом і пошкодження мікротрубочок веретена поділу. Залежно від того, як саме виникла аномалія в кількості хромосом (у результаті мітотичного чи мейотичного поділу), розрізняють соматичну й мейотичну поліплоїдію та анеуплодію.
Одним із механізмів виникнення автополіплоїдних клітин є явище ендореплікації ДНК: клітина проходить декілька циклів реплікації без подальшого виходу в мітоз. Інший механізм - порушення мікротрубочок веретена поділу, що приводить до нерозходження хромосом або хроматид. Поліплоїдію можна викликати і штучним шляхом, застосовуючи речовини - блокатори мітозу (колхіцин, колцемід та ін.). Ці сполуки та їхні аналоги інгібують утворення мікротрубочок веретена поділу, і хромосоми після реплікації не розходяться до полюсів клітини. Крім того, автополіплоїдні клітини можна отримати шляхом блокування не власне поділу ядра (каріокінезу), а поділу цитоплазми (цитокінезу). Два диплоїдних ядра, залишаючись в одній цитоплазмі, при об'єднанні створюють клітину з тетраплоїдним набором хромосом.
Алополіплоїди утворюються штучним шляхом за рахунок міжвидової гібридизації.
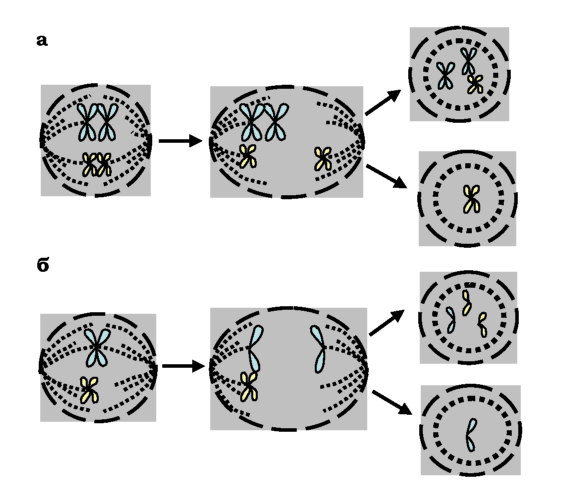
Усі типи анеуплоїдій є результатом нерозходження окремих хромосом (чи хроматид) при поділі клітини - мітозі чи мейозі (рис. 4.8). Найчастіше нерозходження хромосом пов'язане з дефектом центро-мерної ділянки: така хромосома не прикріплюється до веретена поділу й опиняється в одній дочірній клітині разом зі своїм гомологом, інша дочірня клітина виявляється позбавленою однієї хромосоми. Отже, усі форми анеуплодій (моносомія і трисомія, нулісомія і трисомія) можуть бути результатом одного циклу неправильного розподілу хромосом. При мейотичному поділі нерозходження хромосом може відбуватися як у першому, так і в другому поділі.
Поліплоїдні та анеуплоїдні клітини характеризуються порушенням процесів мейозу. Так, при утворенні гамет у поліполоїдів замість бівалентів (див. розділ 1) можуть утворюватися три-, тетра- та уніваленти, що порушує сегрегацію хромосом і викликає появу нових геномних мутацій. Крім того, дисбаланс у кількості хромосом приводить до аномальних мітотичних поділів.
 |
|
Рис. 4.8. Схема утворення анеуплоїдних клітин при першому мейотичному поділі (а) і при другому мейотичному поділі чи мітозі (б) |
Індукція мутацій мутагенними факторами
За своєю природою мутагени поділяють на фізичні, хімічні та біологічні. Коли мутагенний фактор безпосередньо взаємодіє з ДНК і викликає пошкодження, говорять про пряму дію мутагену. Мутаген може не взаємодіяти з ДНК, але запускати каскад процесів, які в кінцевому наслідку приводять до появи пошкоджень (або до інгібування їхньої репарації). У цьому випадку йдеться про опосередковану дію мутагену. Більшість мутагенних факторів, незалежно від їхньої природи, мають як пряму, так і опосередковану дію. Майже всі мутагени є одночасно й канцерогенами, тобто вони здатні стимулювати розвиток пухлин.
Хімічні мутагени за своєю структурою та механізмами дії являють собою дуже різноманітну групу сполук. Хімічні мутагени можуть бути нормальними метаболітами клітини - аутомутагени, або потрапляти в організм (клітину) ззовні - ксенобіотики Найчастіше хімічні мутагени класифікують за хімічною структурою або за типом реакції з ДНК.
Деякі хімічні мутагени є аналогами азотистих основ. Вони за структурою подібні до нормальних основ і можуть використовуватися ферментами, які забезпечують процеси синтезу нуклеїнових кислот. Наприклад, 5-бромурацил є аналогом тиміну і, відповідно, може включатися замість нього в ДНК. Проте, на відміну від тиміну, 5-бромурацил значно легше піддається таутомерізації: у результаті в наступному реплікаційному циклі напроти 5-бромурацилу до ДНК часто включається гуанін.
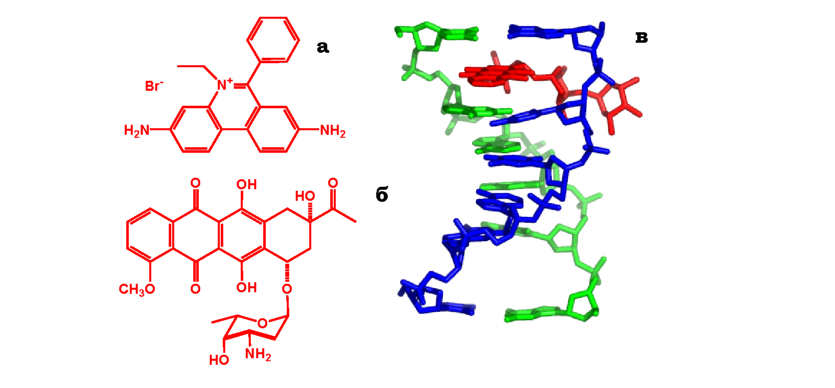
Ароматичні сполуки, здатні до інтеркаляції (вбудовування) між сусідніми парами основ подвійної спіралі ДНК, належать до інтерка-люючих мутагенних чинників. Прикладами є бромистий етидій і дау-номіцин (рис. 4.9, а, б), профлавін, актиноміцин D тощо - планарні молекули, які за розміром наближаються до пурин-піримідинової пари. Вбудовуючись у подвійну спіраль, інтеркалятор удвічі збільшує відстань між сусідніми парами основ (рис. 4.9, в). Під час реплікації відбувається інсерція додаткового нуклеотиду в місці інтеркаляції, що призводить до мутації - зсуву рамки зчитування.
 |
|
Рис. 4.9. Хімічні формули бромистого етидію (а) і дауноміцину (б) і структура комплексу інтеркальованої похідної дауноміцину (червона) з ДНК (в, зображення створено за допомогою програми PyMOL, код структури у Protein Data Bank 1NAB) |
Різноманітні природні та синтетичні речовини, взаємодіючи з ДНК, викликають хімічні модифікації азотистих основ. Скажімо, сильні окисники, такі як азотиста кислота або біхромат калію, ви-
кликають окисне дезамінування основ. Алкілуючі агенти (нітрозо-метилсечовина, тіофосфамід та інші) приєднують алкільні радикали як екзоциклічні групи азотистих основ.
Деякі хімічно інертні молекули, що потрапляють в організм, набувають мутагенних властивостей тільки внаслідок їхніх метаболічних перетворень - метаболічної активації. Такі хімічні речовини називають промутагенами - це, наприклад, гідрофобні поліциклічні ароматичні вуглеводні, афлотоксин (токсин, що синтезується багатьма мукоровими цвілевими грибами).
Узагалі, пошкодження ДНК хімічними речовинами далеко не завжди спричиняється безпосередньою взаємодією мутагену з азотистими основами чи цукрофосфатним остовом. Часто хімічні речовини вступають у складні внутрішньоклітинні реакції, що супроводжуються появою вільних радикалів, які пошкоджують ДНК. Генераторами вільних радикалів можуть виступати сильні окисники, солі металів, деякі антибіотики, наприклад блеоміцин. Хімічними мутагенами, які не взаємодіють з ДНК, є також інгібітори ферментів метаболізму нуклеїнових кислот і репарації. Так, сильним інгібітором систем репарації ДНК є алкалоїд кофеїн. Аналог тиміну - 5-фторурацил - сам у ДНК не вбудовується, а є інгібітором ферменту тимідилатсинтетази. Результатом є нестача тимінових нуклеотидів у клітині, що може бути причиною утворення одноланцюгових прогалин при реплікації ДНК.
Фізичні мутагенні фактори представлені електромагнітним випромінюванням із довжиною хвилі менше З00 нм і корпускулярними випромінюваннями. Іонізуюче випромінювання (рентгенівські та у-промені, а- і в-частинки) викликає іонізацію молекул - втрату чи приєднання електронів, унаслідок чого утворюються позитивно чи негативно заряджені радикали компонентів нуклеїнових кислот. Хімічні реакції між цими радикалами викликають руйнування різноманітних ковалентних зв'язків: фосфодіефірних, глікозидних, зв'язків усередині азотистих основ і цукрів. Подібні процеси індукують як точкові, так і хромосомні мутації всіх типів. Крім прямої дії на ДНК, іонізуюче випромінювання індукує появу вільних гідроксильних і пероксидних радикалів, які підсилюють пряму мутагенну дію іонізуючої радіації.
Серед неіонізуючого випромінювання найбільшу мутагенну активність мають ультрафіолетові промені короткохвильового діапазону (100-280 нм). Саме в цій спектральній області знаходяться максимуми поглинання світла азотистими основами. Поглинання енергії ультрафіолетових променів веде до збудження електронів - їхнім переходам на вищі енергетичні рівні, що дає можливість проходженню фотохімічних реакцій між азотистими основами. Найчастіше продуктами таких реакцій є піримідинові димери, наприклад, між сусідніми (по одному ланцюгу) тимінами (див. рис. 1.18). Довгохвильовий максимум поглинання основ припадає на 260 нм: світло з довжиною хвилі понад 280 нм практично не поглинається ДНК. Певна мутагенна активність довгохвильового ультрафіолету пояснюється поглинанням енергії іншими хромофорами (наприклад, деякими коферментами) із подальшою передачею електронного збудження на ДНК. Крім того, поглинута енергія може також бути причиною генерації вільних радикалів деякими молекулами (скажімо, рибофлавіном).
До біологічних мутагенних факторів відносяться віруси, бактерії, паразити. Основною причиною мутагенної активності вірусів (ретро-вірусів і вірусів, що містять ДНК, див. розділ 5) є інтеграція їхніх ДНК у геном хазяїна. Інтеграція в межах кодуючої або регуляторної ділянки гена може викликати мутацію або вплинути на нормальну експресію. Мутацію може спричинити екзогенна ДНК, штучно введена в клітину. Подібний механізм мутагенезу є цілком подібним до ефектів переміщення мобільних елементів (розділ 6). Крім того, причиною збільшення загального рівня мутацій у клітинах, оброблених екзогенною ДНК або заражених вірусами, є "конкуренція" чужорідної ДНК із клітинною за молекулярну машинерію систем репарації.
Зростання рівня мутацій різного типу, що спостерігається в соматичних клітинах організмів, які піддалися вірусним, бактеріальним та іншим інфекціям, носить назву інфекційного мутагенезу. Інфекційний мутагенез пояснюється комплексом процесів, що відбуваються при взаємодії організму хазяїна з інфекційним агентом або паразитом. Причиною мутацій можуть бути, наприклад, токсини, які виділяють бактерії та паразити, токсичні метаболіти й речовини, синтезовані організмом при реакціях імунної відповіді.
НАСЛІДКИ МУТАЦІЙНОЇ МІНЛИВОСТІ
Загальним наслідком мутаційної мінливості є порушення спадкових програм клітин і організмів. Але при цьому мутаційна мінливість спричиняє підвищення біологічного різноманіття: забезпечує появу нових геномних варіантів і, відповідно, генотипових і фенотипових форм. Більшість з утворених варіантів геному є відносно нейтральними. Велика частина новоутворених фенотипів є або нежиттєздатними (прояв летальних мутацій), або мають знижену життєздатність (прояв напівлетальних мутацій). Проте інколи нові варіанти набувають адаптивної переваги. Адаптивні та нейтральні варіанти закріплюються в популяціях - саме вони забезпечують такі явища, як множинний алелізм і генетичний поліморфізм (див. розділи 3 і 8).
Слід зауважити, що точкові мутації приводять до зміни спадкової програми не обов'язково тільки тоді, коли вони відбуваються в кодуючих ділянках геному. Нуклеотидні заміни в некодуючих ділянках можуть впливати на експресію генів: заміни нуклеотидів у 5'- або 3'-кінцевих ділянках мРНК, що не транслюються, можуть впливати на час життя мРНК; заміни в інтронах - на ефективність сплайсингу; у регуляторних ділянках гена - на рівень експресії. Таким чином, не завжди поліморфізм ДНК у некодуючих ділянках є нейтральним.
Говорячи про наслідки мутаційної мінливості, слід відмежувати ті, що стосуються окремого організму (індивідуальні наслідки), від наслідків для популяцій і видів у цілому (еволюційні наслідки). Мутації в соматичних клітинах часто приводять до негативних ефектів для окремих організмів: наприклад, усі типи мутацій, від точкових до ге-номних, можуть бути причиною виникнення злоякісних новоутворень. Але такі мутації не спадкуються в наступних поколіннях нащадків. Генеративні мутації зумовлюють розвиток особин, усі клітини яких будуть нести дану зміну. Типовим проявом таких мутацій є різноманітні спадкові хвороби (див. розділ 7).
Зрозуміло, що для популяцій і груп організмів, а отже, і для видоутворення, найбільш значимими є генеративні мутації. У ході розвитку різних класів хребетних (від круглоротих до ссавців) спостерігається значна кількість хромосомних і геномних перебудов, які в основному представлені транслокаціями, інверсіями та змінами кількості хромосом. Так, людина відрізняється від людиноподібних мавп робертсо-нівською транслокацією (друга хромосома людини є транслокаційною формою двох акроцентричних хромосом мавп, див. розділ 7).
Важливу роль в еволюції рослинного й тваринного світу відіграють геномні мутації (поліплоїдії та анеуплодії). Проте, як правило, відсутність у каріотипі однієї з хромосом гомологічної пари приводить до зниження життєздатності особин, а в деяких випадках має летальні наслідки. Такий ефект можна пояснити або недостатністю певних білкових продуктів (експресуються гени лише однієї хромосоми), або наявністю в хромосомі, яка присутня в анеуплоїдному ядрі, летальних алель-них варіантів певних генів. Слід зазначити, що фенотиповий ефект мо-носомії залежить від того, яка хромосома втрачена. У людини, наприклад, моносомії за статевою Х-хромосомою (синдром Шерешевського -Тернера, див. розділ 7) мають менш несприятливий для організму ефект порівняно з моносоміями за аутосомами, які є летальними. У дрозофіли нежиттєздатними є особини з нестачею четвертої хромосоми, тоді як нестача другої, третьої або Х-хромосоми не має летальних наслідків. У рослин (пшениці, кукурудзи, тютюну) моносомні форми можуть не відрізнятися від нормальних диплоїдних особин або мати незначні відмінності в розмірах окремих частин рослини.
Що стосується наявності в каріотипі додаткової хромосоми, то зниження життєздатності або летальність трисомій загалом характерна для представників тваринного світу. Рослини-трисоміки часто є життєздатними й мають незначні відхилення від нормальних організмів: у дурману Datura stramonium, наприклад, описані життєздатні трисоміки по кожній із 12 пар хромосом.
Поліплоїдія найбільше розповсюджена в рослинному світі. Збільшення наборів хромосом у рослин приводить до збільшення їхньої вегетативної маси та розвитку стійкості до несприятливих умов. Розпо-всюдженість поліплоїдних форм у рослин пов'язана з тим, що для них характерне вегетативне розмноження та самозапилення. Поліплоїдні форми у тварин найчастіше не є життєздатними, оскільки поліплоїдія несумісна зі статевим процесом. Часто поліплоїдія у тварин зумовлює їхню стерильність, особливо це стосується алополіплоїдних форм. Відповідно, поліплоїдні форми є характерними для тварин із партеноге-нетичним розмноженням.
Будь-який організм є відкритою системою: реалізація спадкової програми відбувається не тільки під контролем генотипу, але й під впливом оточуючого середовища. Умови середовища можуть зумовлювати як ступінь прояву спадкової ознаки (експресивність), так і ймовірність прояву ознаки взагалі (пенетрантність). Таким чином, особини з однаковими генотипами, залежно від впливу навколишнього середовища, можуть мати відмінні фенотипи. Така мінливість називається фенотиповою, або модифікаційною. Класичним прикладом фенотипових змін у межах одного організму є різні форми листової пластинки у стрілолиста - залежно від розташування листя в підводній або надводній частині рослини (рис. 4.10). Модифікаційним змінам в основному піддаються кількісні та якісні полігенні ознаки. Наприклад, зріст, вага, схильність до розповсюджених хвороб, тривалість життя значно залежать від умов середовища.

Модифікаційна мінливість зумовлена не змінами послідовностей ДНК, а змінами в експресії генів (від ефективності транскрипції до особливостей перебігу біохімічних реакцій) під впливом факторів навколишнього середовища. Очевидно, що такі зміни не спадкуються, проте межі модифікаційної мінливості (норма реакції) для кожної окремої ознаки повністю визначаються генотипом. Розмах норми реакції є адаптивною ознакою та визначає межі змін умов навколишнього середовища, в яких може існувати особина, тобто окремий генотип.
Результатом модифікаційної мінливості може бути поява організмів, які фенотипово копіюють прояв певного генотипу. Такі фенотипові форми називають фенокопіями. По суті, фенокопії є проявом крайніх варіантів норми реакції. Наприклад, недостатність вітаміну D спричинює розвиток рахіту, який за симптоматикою нічим не відрізняється від спадкового захворювання, зумовленого мутаціями, які приводять до нечутливості до вітаміну D.
До окремого типу модифікаційної мінливості відносять так звані довготривалі модифікації, які можуть спадкуватися протягом декількох поколінь. Відомим прикладом є зміна забарвлення в колорадського жука, яка виявляється після витримування лялечок за високих температур. Змінене забарвлення зберігається для кількох поколінь, а потім повертається до вихідного стану. При цьому спадкування відбувається виключно по материнській лінії: змін у геномі не відбувається, а тривалість модифікації пояснюється накопиченням у цитоплазмі клітин зародкового шляху стабільних мРНК генів теплового шоку, які з цитоплазмою яйцеклітини передаються нащадкам. Зрозуміло, що через декілька поколінь відбудеться "розведення" цих мРНК: їхня кількість стане врешті-решт недостатньою для прояву зміненого забарвлення.
Контрольні запитання і завдання
1. Дайте визначення терміну "мінливість". Які існують типи мінливості?
2. Чим зумовлена спадкова мінливість?
3. Що таке мутації? Чим соматичні мутації відрізняються від генеративних.
4. Опишіть типи точкових мутацій.
5. Опишіть типи хромосомних і геномних мутацій.
6. Що є джерелом мутаційних змін?
7. Які пошкодження ДНК виникають у процесі життєдіяльності клітин і внаслідок помилок реплікації та репарації?
8. Опишіть механізми виникнення хромосомних і геномних мутацій.
9. Дайте визначення поняття "мутагенний фактор". Що таке пряма та опосередкована дія мутагену?
10. Які типи хімічних мутагенів ви знаєте. Опишіть механізми їхньої дії. Що таке промутагени?
11. Поясніть механізми мутагенної дії фізичних факторів.
12. Які мутагенні чинники можна віднести до факторів біологічної природи? Які механізми їхньої дії?
13. Які наслідки мутаційної мінливості є негативними? Позитивними?
14. Що таке модифікаційна мінливість? Які основні причини її прояву? Чим модифікаційна мінливість відрізняється від мутаційної?
15. Що таке фенокопії?
16. Що таке довготривалі модифікації? Поясніть механізми їхнього утворення.
Автор: admin от 18-07-2013, 16:54, Переглядів: 17843